

Bacterial wilt caused by Ralstonia pseudosolanacearum infection has led to drastic loss of tomato fruit worldwide. Planting tomato plants harbouring genetic resistance to bacterial wilt have been suggested for desirable and durable tomato productions (Jung et al., 2014; Kim et al., 2016; Lee et al., 2015). However, the cultivar resistance of tomato plants to bacterial wilt can be overcome by newly emerging pathogenic bacterial strains and changing climates such as elevated temperatures and ambient CO2 concentration (Lee et al., 2011, 2015; Mew and Ho, 1977; Shin and Yun, 2010). Limited available chemical pesticides and concerns on environmental contamination and human health provoked exploring novel promising abiotic and biotic agents for efficiently managing the tomato bacterial wilt. Application of chemical plant defence activators and antagonistic microorganisms have reduced the tomato bacterial wilt (Hong et al., 2011; Jogaiah et al., 2013; Tan et al., 2013). In our previous studies, various approaches to control the tomato bacterial wilt using chemical pesticides and eco-friendly compounds have been tried so far. Copper-based chemical pesticides delayed bacterial wilt development in tomato plants (Lee et al., 2012). Soil-drenching diluted clove oil solution and hydrogen peroxide-nitric oxide mixed aqueous solution effectively attenuated symptom development of the tomato bacterial wilts as well (Lee et al., 2012; Hong et al., 2013). Recently, thiamine (vitamin B1) was also suggested to slightly ameliorate bacterial wilt in detached tomato leaves (Hong et al., 2016). We have been trying to find more efficient and convenient control methods for eco-friendly and integrated tomato bacterial wilt management together with the several agents suggested previously.
Peroxyacetic acid and peroxyacetic acid-contained mixtures have been introduced for eradicating hazardous microbes in postharvest periods of horticultural crops. Human pathogenic Escherichia coli O157:H7 could be removed from the apple fruit surface by peroxyacetic acid treatment depending on the dosage and washing time applied (Wisniewsky et al., 2000). Immersing strawberry fruits in peroxyacetic acid solution for 2 min has led to reduction in population of food poisoning bacteria E. coli and Salmonella enterica on the fruit surface (Gurtler et al., 2014). Spraying peroxyacetic acid onto flowers and developing immature fruits of strawberry plants prior to fruit harvest delayed fruit decay caused by Botrytis cinerea infection during the postharvest storage period (Narciso et al., 2007). S. enterica on the tomato fruit surface was efficiently eliminated by peroxyacetic acid treatment (Chang and Schneider, 2012; José and Vanetti, 2012). Commercial peroxyacetic acid mixtures containing different composition of peroxyacetic acid, hydrogen peroxide and acetic acid have been available worldwide, and usually they have been suggested as fruit sanitizers and disinfectants after the vegetables and fruits have been harvested. These findings indicated that peroxyacetic acid itself or mixtures containing peroxyacetic acid as a component may be also applied to the horticultural crops to reduce various troublesome diseases during cultivation in the field unless it causes adverse effect on vegetative and reproductive plant growths.
It is barely demonstrated that application of peroxyacetic acid-mediated crop protection against pathogen infections in open-fields or protected greenhouses during plant cultures including tomato production. Vines et al. (2003) tried to apply peroxyacetic acid on tomato plants growing in a hydroponic culture system, indicating that peroxyacetic acid treatment for crop protection during the plant production should be carefully considered because of the phytotoxic effect caused by a high dosage of peroxyacetic acid. Successful application of a commercial peroxyacetic acid-associated agent was reported only in floriculture for foliar nematode control, which evidenced a sustainable way to use peroxyacetic acid in eco-friendly disease management during crop cultures (Jagdale and Grewal, 2002).
In this study, possibility of peroxyacetic acid mixture Perosan (peroxyacetic acid 4%, H2O2 27%), commercially available as an agricultural disinfectant, was evaluated whether or not it could reduce bacterial wilt in tomato plants. In vitro bacterial growth of R. pseudosolanacearum influenced by the peroxyacetic acid mixture treatment was investigated. The Perosan dosage-dependent responses of the detached tomato leaves and whole tomato seedlings were uncovered to suggest safe usage of the peroxyacetic acid mixture without phytotoxic effect on the tomato plants. Delayed bacterial wilt occurred in the detached tomato leaves and whole tomato seedlings treated with the peroxyacetic acid mixture were also demonstrated.
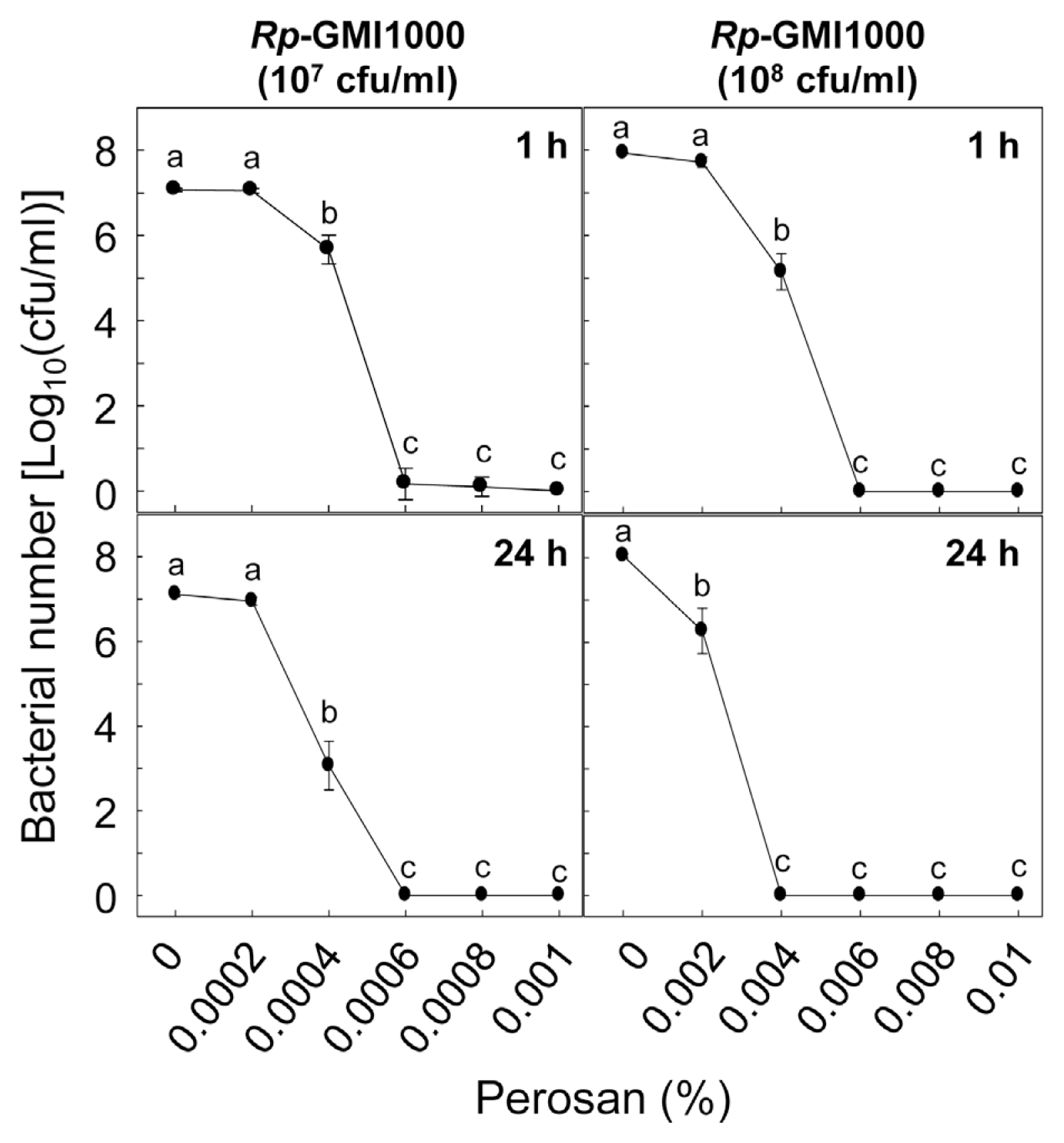
Effective dosages of the peroxyacetic acid mixture for suppressing higher population of bacteria was evaluated in vitro (Fig. 1). Different doses of the peroxyacetic acid mixture were applied to the two bacterial suspensions of R. pseudosolanacearum (107 and 108 cfu/ml) in microtubes for 1 and 24 h at 30°C. The peroxyacetic acid mixture-treated bacterial cells were inoculated on CPG agar media supplemented with 2,3,5-triphenyltetrazolium chloride (TTC) and incubated for 2 days at 30°C (Lee et al., 2012). Relatively small numbers of the bacterial colonies were formed on the CPG agar media by the Perosan treatment. At least 0.0004% of the peroxyacetic acid mixture began to decrease survival of 107 cfu/ml of bacteria, and more than 0.0006% of peroxyacetic acid mixture completely killed all bacterial cells within 1 h. Incubation for 24 h with 0.0004% of Perosan more suppressed the bacterial growth. To kill 108 cfu/ml of the bacteria completely, relatively higher Perosan (0.006%) was required. Further incubation for 24 h with 0.004% of Perosan completely suppressed the bacterial growth. It can be suggested that direct treatment with Perosan was efficient to control bacterial propagation and dissemination in soil environment highly populated by the R. pseudosolanacearum.
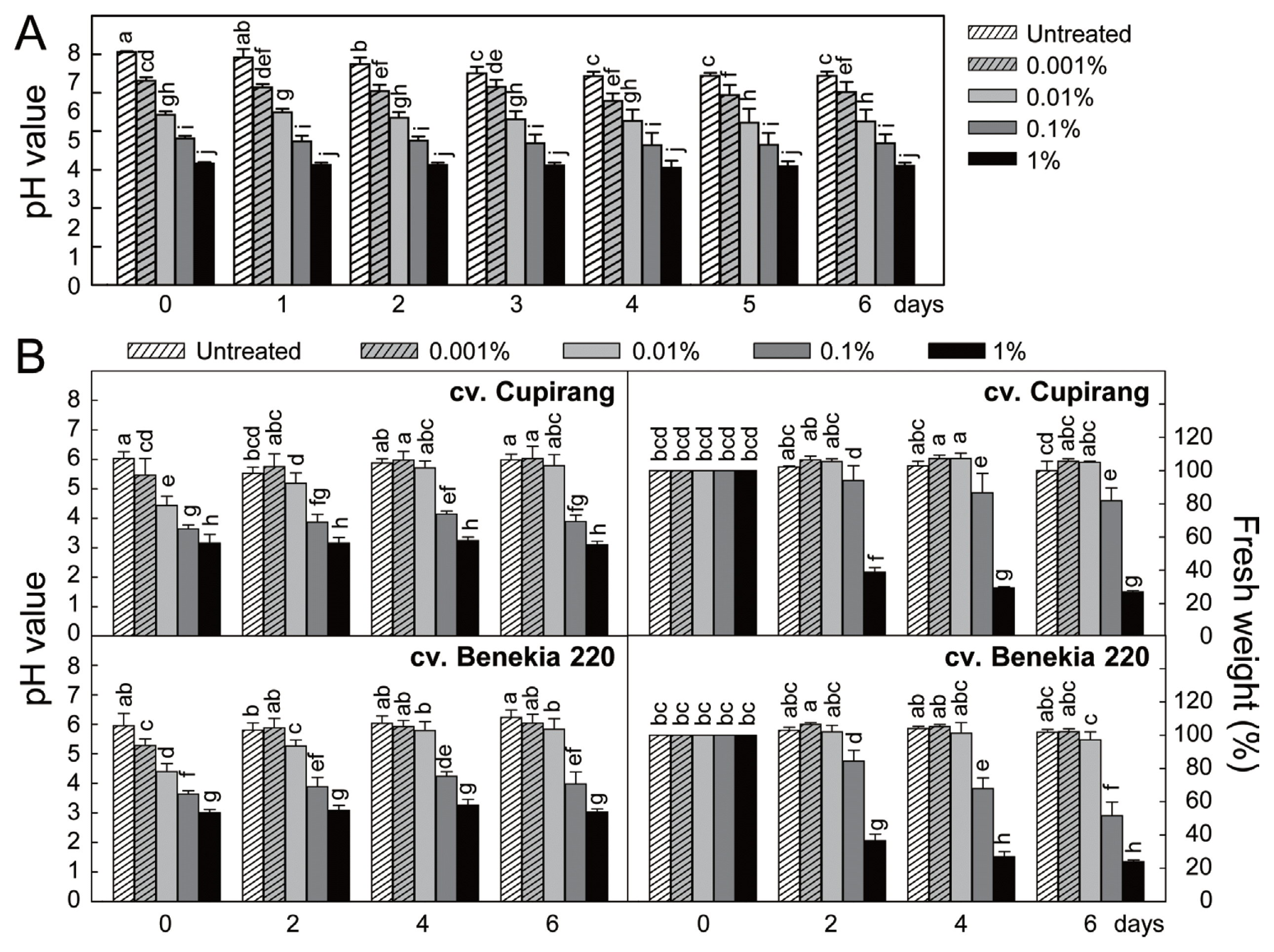
Phytotoxicity of the Perosan was investigated in two susceptible tomato cultivars (Cupirang and Benekia 220) (Fig. 2). Before being applied to tomato plants, different concentrations of Perosan solution were investigated in light of pH change because Perosan concentrate is highly acidic, and this raised concerns about adverse effects on the plant growing (Fig. 2A). Treatment with increasing concentrations of Perosan (0, 0.001, 0.01 and 1%) has lowered the pH in the aqueous solutions. Immediate drops in pH mediated by Perosan treatments did not seem to be restored by 6 days after treatments. Temporal changes in pH of the Perosan-treated solution and in fresh weight (FW) of the detached tomato leaves were investigated (Fig. 2B). Tomato seedlings were grown in a walk-in growth chamber to the five-leaf stage under controlled environmental conditions previously described (Hong et al., 2016), and the third true leaves were detached and placed in 10 ml of different concentrations (0, 0.001, 0.01, 0.1 and 1%) of Perosan solution by using the petiole dipping method (Hong et al., 2016). Treatment with 0.001 and 0.01% of Perosan decreased the pH prior to the petiole dipping, but pH was recovered overtime after the dipping petioles of cv. Cupirang. The low pH mediated by 0.001% of Perosan was recovered overtime by the dipping petioles of cv. Benekia 220 as well, but the decreased pH by 0.01% of Perosan still slightly remained low. Distinctly low pH caused by 0.1 and 1% of Perosan was not compromised by petiole dipping of two tomato cultivars. It is supposed that the tomato leaf petiole exudates may be not enough to recover low pH caused by higher Perosan concentrations 0.1 and 1%. Relatively high concentrations of Perosan led to reduced leaf growth and tissue damage in the detached leaves of both tomato cultivars, shown as significantly decreased FW of the detached leaves. It can be suggested that more than 0.1% of Perosan should be avoided not to cause phytotoxic effect in the tomato leaf tissues during hydroponic cultures.
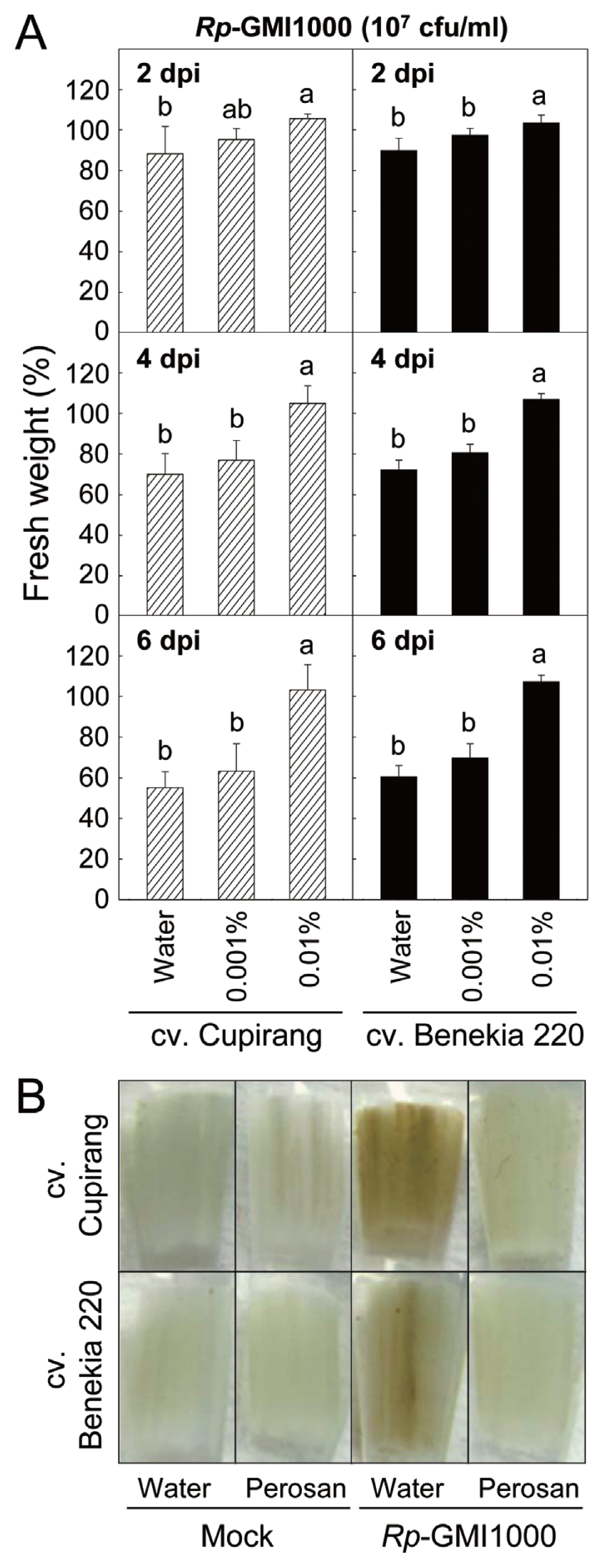
Crop protection efficacies of the Perosan treatment were evaluated in detached leaves of two different tomato cultivars (Cupirang and Benekia 220) inoculated with R. pseudosolanacearum (Fig. 3). The third true leaves of tomato seedlings at five-leaf stage were detached for the bacterial inoculation (107 cfu/ml) and relative FW of the leaves was expressed as the effect of a wilting symptom development for 6 days described in our previous study (Hong et al., 2016). Wilting developments of the detached tomato leaves treated with the Perosan were retarded in both cultivars in a dose-dependent manner (Fig. 3A). Low concentration (0.001%) of Perosan could not protect the detached tomato leaves, but relatively high concentration (0.01%) of Perosan effectively suppressed bacterial wilt of the detached tomato leaves of the two cultivars without any visible phytotoxic effect. No significant difference between the two tomato cultivars was found in the disease-suppressing activities of the peroxyacetic acid mixture It was interesting that direct incubation of the bacterial suspension (107 cfu/ml) with 0.001% of the Perosan for 1 h in the microtubes without dipping the detached tomato leaf petiole completely suppressed the bacterial growth in Fig. 1, but the concentration was not reducing bacterial wilt of the tomato leaves inoculated with the same inoculum dose of the R. pseudosolanacearum in Fig. 3. Petiole exudates from the detached tomato leaves may inhibit Perosan-triggered bacterial cell death or increase bacterial proliferation at an early colonization step. It was supported by the facts that R. solanacearum showed different chemotactic behaviour to various sugars, organic acids and amino acids (Yao and Allen, 2006). Root exudates from non-host rice plants were less attractive to the R. solanacearum compared to host tomato plants. Oxalic acid, as an inducer of chemotactic response and biofilm formation of R. solanacearum, was highly accumulated in root exudates from bacterial wilt-susceptible tobacco cultivar compared to root exudates of resistant cultivar (Wu et al., 2015). Recovered low pH by tomato petiole dipping may also attribute to decreased protective effect of Perosan as well. Brown-coloured petiole severely damaged by R. pseudosolanacearum infection was comparable with normal petiole tissue inoculated with R. pseudosolanacearum with 0.01% Perosan (Fig. 3B).
Perosan was also applied to soil mixture in pots in which four-leaf stage tomato seedlings were raised to uncover phytotoxicity during the plant growth in soil environments (Fig. 4). By drenching 10 ml of different concentrations (0, 0.5, 1, 2, and 5%) of Perosan into the pots containing commercial soil mixtures, tomato seedlings have shown different phytotoxic responses depending on Perosan concentrations applied (Fig. 4A, 4B). The highest dose (5%) of Perosan solution led to drastic tissue collapse at the lower part of the tomato stems in both cultivars. No distinct adverse effect on growth of the two tomato cultivars was found by less than 2% Perosan treatment, which was quite a high concentration of Perosan to mediate phytotoxic effect on the detached tomato leaves in Fig. 2B. Perosan applied by the drenching method may be hindered for direct contact of tomato roots in the soil environment. Tomato seedlings hydroponically grown supplemented with another peroxyacetic acid mixture Proxitane 0510® (Solvay, Ltd., Brussels, Belgium) (peroxyacetic acid 5%, H2O2 18%, acetic acid 15%) exhibited much sensitivity to the peroxyacetic acid mixture and showed concentration-dependent tissue damages (Vines et al., 2003). Treatment with at least 0.01% of Proxitane 0510® distinctly reduced tomato plant growth for a 1-month hydroponic culture. The tomato tissue damage by the peroxyacetic acid mixture may not be caused by H2O2. An H2O2 dose (1.35%) in 5% Perosan was much lower than concentration used in our previous study demonstrating that soil drenching 10 ml of 200 mM H2O2 (19.72%) did not cause any phytotoxic effect on tomato growth (Hong et al., 2013). Perosan-mediated damage of tomato plants in the present study could be due to low pH conditions and/or oxidative stresses due to peroxyacetic acid included in Perosan.
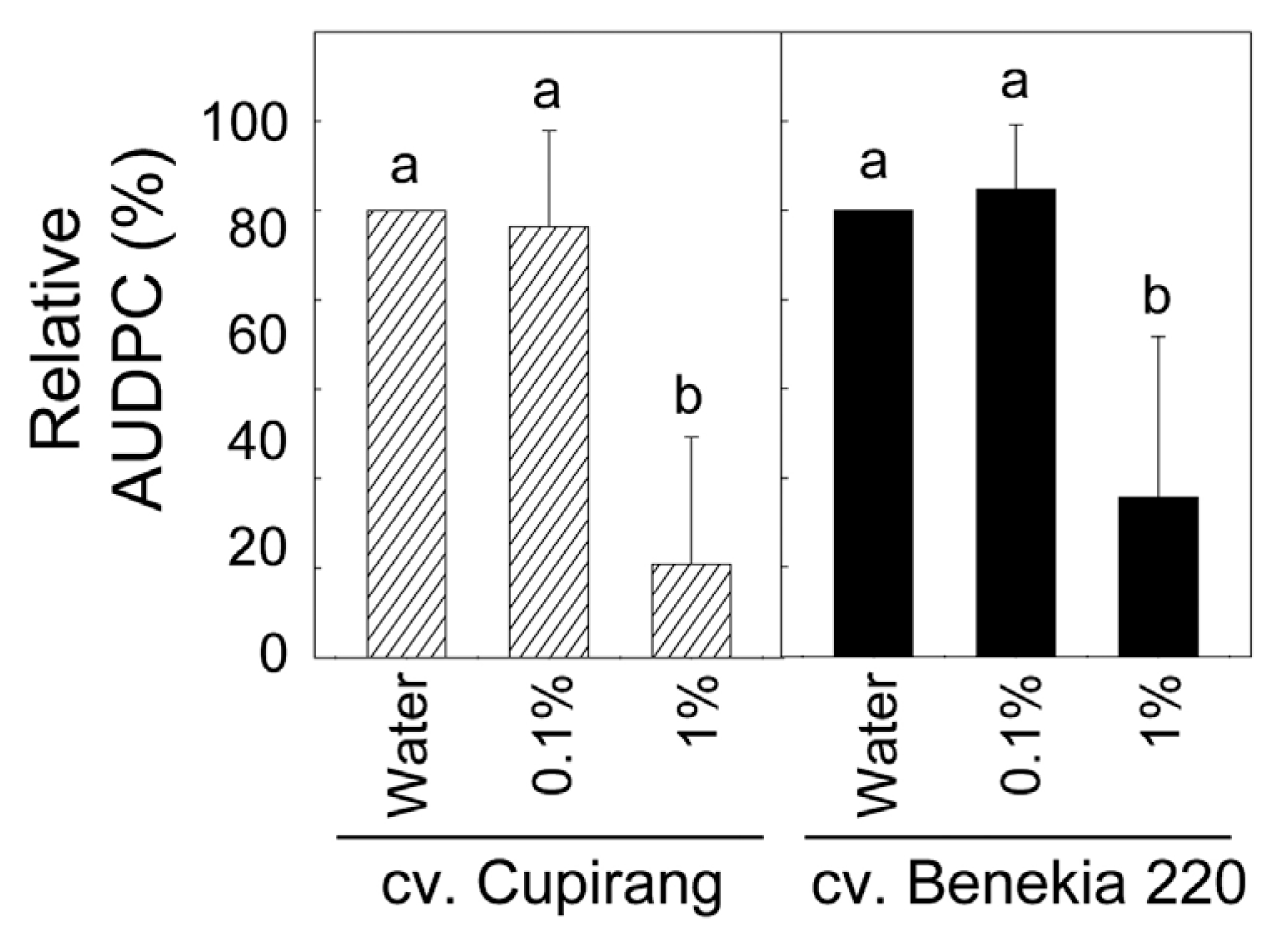
Perosan-mediated disease control of bacterial wilt was also evaluated in the four-week-old tomato seedlings grown in pots and shown as relative areas under disease progress curve (AUDPC) assessed (Fig. 5). Ten ml of R. pseudosolanacearum suspension (108 cfu/ml) was applied to the basal part of the tomato stem by soil-drenching. Distilled water (as an untreated control) and two concentrations of Perosan (0.1 and 1%) were prepared, and 10 ml of each solution was poured to the bacteria-inoculated areas 1 h after the bacterial inoculation. Disease severity of bacterial wilt of tomato was evaluated for 12 days after bacterial inoculation based on the disease scale (0-4 scale) previously described (Lee et al., 2012). One% of Perosan solution led to significantly decreased relative AUDPC in the two tomato cultivars, which suggest that soil-drenching 1% of Perosan can be applied nearby basal stem part of tomato plants to reduce bacterial wilt. However, disease protection efficiency of the Perosan was relatively lower than that derived from 0.01% clove oil application evaluated in our previous study (Lee et al., 2012). To improve Perosan-mediated control of the tomato bacterial wilt, increases in frequency of the application may be considered.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






