

In response to ambient conditions, microbes including pathogenic bacteria, regulate the expression of adaptive genes or gene sets through exchanges of small cell-cell signaling molecules. One of these communication factors that was first described in the phytopathogenic bacterium Xanthomonas campestris pv. campestris (Xcc) has been named as a diffusible signal factor, DSF (Tang et al., 1991). The Xcc genome includes a gene cluster (named as regulation of pathogenicity factor [rpf ] genes) that is involved in the synthesis, detection, and signal transduction of DSF. The rpf cluster contains at least 8 genes, rpfA-H. RpfB and RpfF proteins mediate DSF production, and rpfC and rpfG encode a histidine kinase (HK) and a response regulator (RR), respectively, which together form a two-component regulatory system (TCS) (Barber et al., 1997; He et al., 2007; Slater et al., 2000). The DSF signals mediated by the RpfC/RpfG TCS regulate the expression of genes involved in virulence activities such as production of extracellular enzymes (endoglucanase and protease) and the extracellular polysaccharide (EPS) xanthan (Barber et al., 1997; Fouhy et al., 2006; Slater et al., 2000; Tang et al., 1991). It has been known that these rpf-genes are well-conserved in all xanthomonads and Xylella fastidiosa (Xf) (Chatterjee et al., 2008; Lee et al., 2006).
DSF is often regarded as a quorum-sensing (QS) molecule that allows bacteria to assess its population density (von Bodman et al., 2003). Gram-positive bacteria typically produce peptides (Nagao et al., 2006) as QS signals, and most of Gram-negative bacteria produce homoserine lactone derivatives (de Kievit and Iglewski, 2000) with the exception of activator of XA21-mediated immunity (Ax21), a recently discovered small QS protein in Xanthomonas oryzae pv. oryzae (Xoo) (Han et al., 2011). When the extracellular concentration of a secreted QS signal rises above a specific threshold, the bacterium can recognize the signal, which in turn regulates the expression of genes involved in diverse group behaviors, such as swarming motility, biofilm formation, cell division, stress survival, and production of virulence factors (Fuqua et al., 2001; Whitehead et al., 2001; Withers et al., 2001).
Xoo, a pathogenic Gram-negative bacterium, invades rice leaves through natural pores such as hydathodes (Shen and Ronald, 2002) to cause bacterial leaf blight (BLB), a disease that significantly reduces rice yield in many countries (Cheatham et al., 2009). With genome sequencing completed in a few strains of the bacterium (Lee et al., 2005; Ochiai et al., 2005; Salzberg et al., 2008), information is available to study the bacterial pathogenicity genes. In a previous investigation of the function of rpf genes, we showed that mutations in rpfB, rpfC, rpfF, and rpfG reduce the virulence and motility in Xoo and decrease the expression of genes involved in the production of EPS, lipopolysaccharide (LPS), phytase, xylanase, and lipases (Jeong et al., 2008). However, a role for the rpfE gene, which encodes a regulatory protein (Lee et al., 2006), still remains elusive. Like other rpf- genes, rpfE is well-conserved in all xanthomonads and Xf (Lee et al., 2006), but a characterization of rpfE, albeit brief, was published only for Xcc (Dow et al., 2000). In the present study, we used a Xoo rpfE knock-out mutant strain to clarify the function of rpfE in Xoo virulence and the rpf system. Our findings suggest that rpfE influences Xoo pathogenicity by regulating the production of virulence factors without affecting DSF production.
Material and Methods
Bacterial strains and culture conditions
The bacterial strains and plasmids used in this study are listed in Table 2. Xoo strains were grown in rich media, e.g., nutrient agar (NA) and peptone sucrose agar (PSA), and in minimal media, M9 (Miller, 1972). Escherichia coli strains were cultured in Luria-Bertani (LB) medium. Antibiotics were used at the following concentrations: kanamycin (Km), 50 ╬╝g/ml; ampicillin (Amp), 100 ╬╝g/ml; cephalexin (Cep), 100 ╬╝g/ml; and gentamicin (Gm), 10 ╬╝g/ml.
For growth rate assay, Xoo WT and rpfE-and rpfC-mutant strains were incubated at 28┬║C in M9 minimal medium containing 50 mM glucose, xylose, or sucrose, with shaking at 200 rpm. At indicated time points, 100-╬╝L aliquots of each culture were spread on PSA medium, and the colony forming units (CFUs) were counted after incubation at 28┬║C for 3 days.
Construction of mutant and complementation strains, and molecular techniques
All primers for the construction of the rpfE knockout mutant and rpfE complementation strains (Table 3) were designed on the basis of the Xoo KACC10331 genomic sequence (GenBank accession number AE013598) (Lee et al., 2005). The rpfE gene was amplified from Xoo genomic DNA and cloned into a TA-cloning vector (RBC Bioscience Corp., Taipei, Taiwan). Successful cloning was confirmed by sequencing and Xoo rpfE was subsequently cloned into the pUC19 vector using restriction enzymes (BamHI/HindIII) (Invitrogen Corporation, Carlsbad, CA, USA). The EZ-Tn5TM<KAN-2> TransposomeŌäó insertional kit (Epicentre Biotechnology, Madison, WI, USA) was used to insert Tn5 into rpfE:: pUC19. A clone containing the Tn5 insertion in the middle of the rpfE gene was selected by restriction enzyme screening, and the location of insertion was confirmed by nucleotide sequencing analysis (Microgen Inc., Seoul, South Korea). The selected plasmid was introduced into Xoo KACC10859 by electroporation to generate the rpfE knockout strain. Transformants with homologous recombination were isolated on selection medium containing Km and Amp and confirmed using PCR-based analysis. To determine if phenotypic changes in the knockout mutant strain occurred through a polar effect, a complementation construct was generated using an expression vector, pML122 (Labes et al., 1990), as previously described (Cho et al., 2011; Labes et al., 1990). The rpfE gene containing no promoter region amplified by PCR using the primers rpfE-com-F/R (Table 3) was inserted into TA vectors. The cloned fragment was verified with sequencing analysis and inserted into pML122 using restriction enzymes (XhoI/BamHI), and the plasmid carrying Xoo rpfE was introduced into the knockout mutant strain by electroporation. The construct was also verified by restriction enzyme digestion and PCR-based analysis before Xoo transformation.
In this study, total DNA was extracted from Xoo cells using ExgeneŌäó cell SV (GeneAll Biotechnology Co. Ltd., Seoul, South Korea), plasmids were purified using the plasmid mini-prep ExprepŌäó plasmid SV (GeneAll Biotechnology Co. Ltd., Seoul, South Korea), and PCR was performed with Ex TaqŌäó (Takara Biotechnology Inc., Tokyo, Japan). The Gene Pulser XcellŌäó system (Bio-Rad Laboratories, Hercules, CA, USA) was used to transform Xoo with the following settings: voltage pulse, 3.0 kV; capacitance, 25 ╬╝F; and resistance, 200 ╬®.
Enzymatic activity and swarming assays
Cellulase activities were assayed as described (Chatterjee et al., 1995). The rpfE mutant was incubated in Nutrient Broth (NB) for 72 h, and the optical density of the cultures was then adjusted to 1.0 at 600 nm. The culture supernatant was added to cellulase assay medium and incubated for 24 h at 28┬░C. The cellulase detection plate was stained for 10 min with 0.1% Congo Red and washed 3 times with 1 M NaCl until it showed a clear zone. Xylanase activity and EPS dry weight were determined as previously described (Jeong, 2008). The rpfE mutant strain was cultured in NB for 72 h and the optical density of the cultures was then adjusted to 1.0 at 600 nm.
To determine xylanase activity, 1 ml of culture supernatant was incubated with 480 ╬╝l of xylanase assay solution for 30 min at 28┬░C. To this mixture, 960 ╬╝l of absolute ethanol was added and the reaction was incubated at room temperature for 40 min. The mixture was centrifuged at 10,000 ├Ś g for 10 min, and the optical density of the supernatant was measured at 590 nm. To measure EPS dry weight, 1.0% KCl (w/v) was added to the culture supernatant. Two volumes of absolute ethanol were then added to the tubes, and the mixture was incubated at ŌłÆ20┬░C overnight. The mixture was centrifuged at 10,000 ├Śg, the EPS pellets were dried for 12 h at 55┬░C and the dry weight of the pellet was measured.
The swarming test was performed on peptone sucrose soft agar containing 0.3% agar as previously described (Ryan et al., 2007; Shen et al., 2001). All virulence-related phenotype assays were independently repeated 3 times.
Gene expression analysis by quantitative RT-PCR
The total RNA was extracted from Xoo KACC10859, the rpfE mutant strain, and the complementation strain using RiboPureŌäó Kits (Invitrogen Corporation) after culture for 2 days at 28┬░C. A QuantiTect┬« Reverse Transcription Kit (QIAGEN, D├╝sseldorf, Germany) was used to synthesize cDNA, which was then subjected to quantitative real-time PCR (qRT-PCR) that was performed with SYBR┬« Premix Ex TaqŌäó (Takara Bio, Inc.) using a Smart Cycle┬«II system (Takara Bio, Inc.) and SeeAMPŌäó (Seegene, Inc.). Primers were designed on the basis of Xoo KACC10331 genome sequence in GenBank (Table 2). qRT-PCR was carried out as previously described (Jeong et al., 2008) with an initial denaturation step at 95┬░C for 30 sec, followed by 45 cycles of denaturation at 95┬░C for 5 sec, annealing at 56┬░C for 15 sec, and extension at 72┬░C for 20 sec. The quantity of the synthesized target mRNA was calculated using 16S rRNA as an internal control with QubitŌäó ver. 1.01 (Invitrogen Corp.), and the relative expression of each gene was calculated using the Smart-Cycle┬« 3.0 software and Seegene viewer.
Thin layer chromatography analysis of DSF production
Each rpf-mutant strain and the Xoo WT strain were incubated in NB medium for 72 h at 28┬║C. To remove bacterial cells, the cultures (500 ml) were centrifuged at 5,000 ├Ś g for 20 min at 4┬║C and filtered through a 0.45-╬╝m membrane. To extract DSF, an equal volume of ethyl acetate (EA, DaeJung Chemicals, Gyeonggi, Korea) was added to the supernatant and the mixture was incubated for more than 1 h with shaking at room temperature (He et al., 2010). The ethyl acetate fractions were separated and concentrated by rotary evaporator (Eyela, N-INW, Tokyo Rikakikai Co., Ltd., Japan). The residues were dissolved in 3 mL of CHCl3 and concentrated to a small volume by vacuum centrifugation (SAVANT SpeedVac┬« DNA120, Thermo Scientific, Vantaa, Finland). The samples were spotted on silica plates (TLC Silica gel 60 RP-18 F254s, Merck Korea, Seoul, Korea) with DSF standard (cis-11-methyl-2-dodecenoic acid, Cayman Chemical Company, Ann Arbor, MI, USA), and then developed with 2-butanol: ethanol:water = 2:1:1 (BEW). Separated spots were detected under UV light at 254 nm and extracted in CHCl3.
Inoculation experiment and enumeration of the bacteria in the rice leaf
Xoo strains were inoculated to test virulence ability using the scissor-clip method (Tang et al., 1996). Xoo strains were grown in NB and PSB to optical densities of 0.1 at 600 nm for the inoculation, and the susceptible rice cultivar Milyang 23 or Dongjin 1 was used. Lesion lengths were measured at 14 days after inoculation for end point measurement or measured daily using digital vernier calipers (CD-15CP, Mitutoyo Corporation, Kanagawa, Japan).
For measurement of population change of the pathogen in the inoculated rice leaf, top 10 cm of the inoculated rice leaf were cut by 2 cm and numbered 1st to 5th from the top of the inoculation cut. Each leaflet was cut into small pieces with razor blade and macerated in sterilized water with microfuge tube and plastic pestle. The macerates were vortexed for 20 min and the suspension was plated on NA containing cephalexin, 10 ╬╝g/ml, after the suspension of the macerate was diluted.
Results
Mutation of the rpfE gene in Xoo reduces virulence
The rpfE and flanking region was cloned and sequenced in X. campestris first (Dow et al., 2000; GeneBank AJ2455997.1). The rpfE and its flanking genes, and gene order and orientation were well conserved in X. oryzae pv. oryzae KACC10331 whose genome was sequenced completely (Lee et al., 2005; GeneBank AE013598.1). The 84% of nucleotide sequence and 87 % of amino acid sequence were identity between two genes. The 14 nucleotides were more in rpfExoo at just upstream of 1st codon, which is a major gap between two genes showed by BLAST.
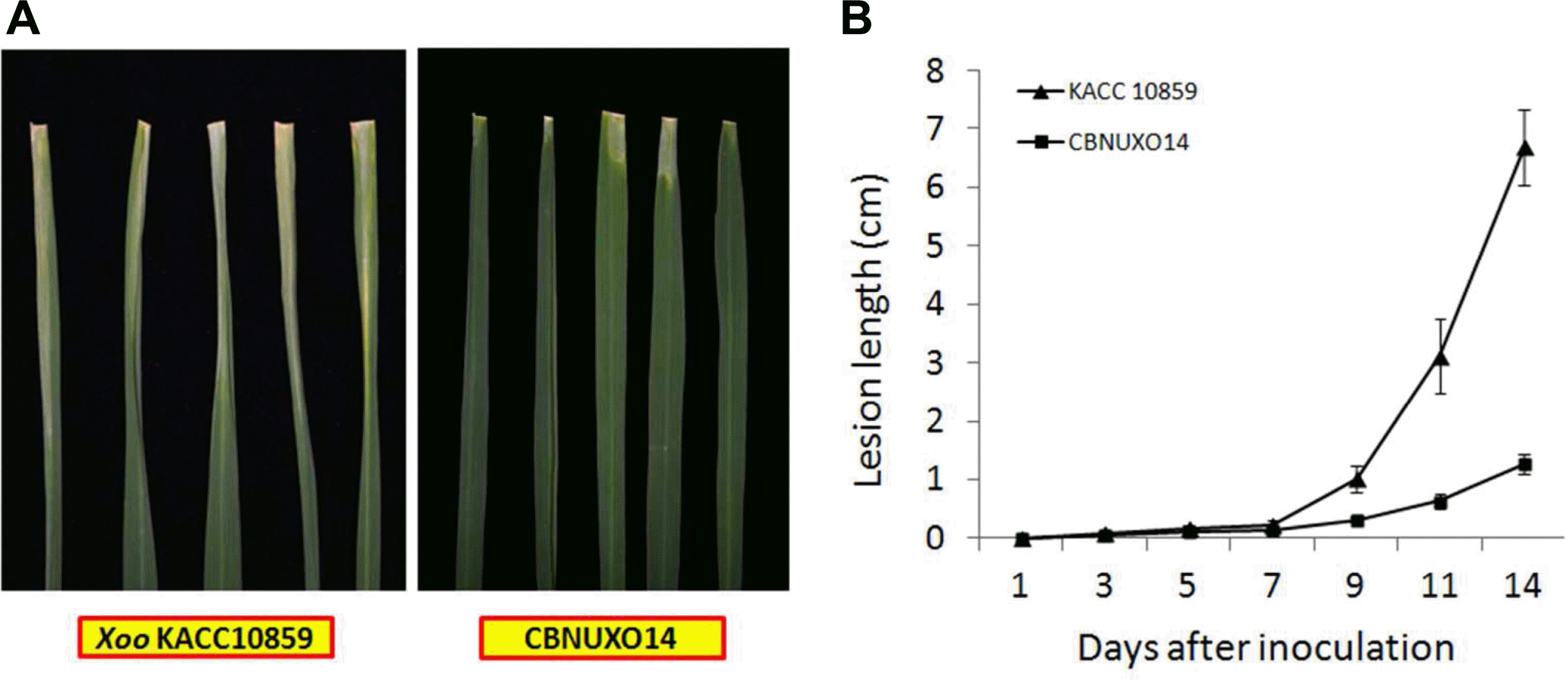
To generate rpfE knock-out mutant, Tn5 was inserted in Xoo rpfE cloned into the pUC19 with EZ-Tn5TM<KAN-2> TransposomeŌäó insertional kit. The sequence of the Tn5 inserted DNA showed that Tn5 was inserted between 595th and 596th nucleotide of Xoo rpfE. By marker exchange, an rpfE-deficient strain (CBNUXO14) was generated. To test the virulence, 6-week-old rice plants (cv. Milyang 23) were inoculated with the mutant (CBNUXO14), a complementation strain (CBNUXO15) and wild-type (WT) KACC10859 strains, and the lesion lengths were monitored for 2 weeks. Lesion lengths were significantly reduced in rice leaves inoculated with the CBNUXO14 strain (8.5 ┬▒ 0.5 cm) compared to leaves inoculated with the WT KACC10859 (28.5 ┬▒ 1.5 cm) and the complementation strain CBNUXO15 (27.9 ┬▒ 1.1 cm) (Table 1). This result indicated that loss of the rpfE gene function substantially reduced the virulence of Xoo.
The disease progress in the inoculated rice leaf was analyzed by time-course monitoring the lesion length and bacterial population using Dongjin 1 cultivar. The lesion length was enlarged rapidly after 7 days post inoculation in wild-type inoculated leaves while the extent and enlargement of lesion was much less in the rpfE-mutant inoculated leaves (Fig. 1). The mutant population (CBNUXO14) was about 10-50 times lower than the wild type population in first 3 days in the inoculation leaflet (Fig. 2, 1st). The rpfE mutant did not build up in the 3rd-5th leaflets, the lower parts of the inoculation leaf, at 7 or 10 days post inoculation (Fig. 2). This result suggests the rpfE mutant did not move to downward from the inoculation point efficiently.
Inactivation of the rpfE gene reduces production of virulence-related factors
To determine whether the production of virulence factors may be affected in the rpfE mutant, extracellular enzyme (cellulase and xylanase) activities and EPS levels were assayed in the CBNUXO14 and CBNUXO15 strains. In the CBNUXO14 strain, cellulase activity was significantly reduced to about 85% of that of the WT KACC10859 strain (Table 1). The EPS dry weight was also reduced to about 40% of that of the WT KACC10859 dry weight (Table 1). The reduction of cellulase activity and EPS production were completely restored by complementation of Xoo rpfE with pML122 (Table 1). However, xylanase activity of the CBNUXO14 strain did not differ from that of the WT KACC10859 strain (Table 1). These results demonstrate that inactivation of rpfE induced reduction of cellulase activity and EPS production in Xoo.
To test the hypothesis that reduction of cellulase activity in the CBNUXO14 strain is dependent on DSF signaling mediated by RpfC/RpfG TCS, cellulase activity in rpfC and rpfG mutant strains was also investigated (Fig. 3). Although reduction of cellulase activity of these mutant strains was observed, it remained more than 50% of that of the WT strain in both mutant strains (Fig. 3).
Another virulence factor, group motility was tested in the rpfE mutant strain. CBNUXO14 had significantly reduced motility in a swarming assay, whereas motility of the complementation strain CBNUXO15 was similar to that of the WT strain (Table 1).
Expression of virulence genes is reduced in the rpfE mutant strain
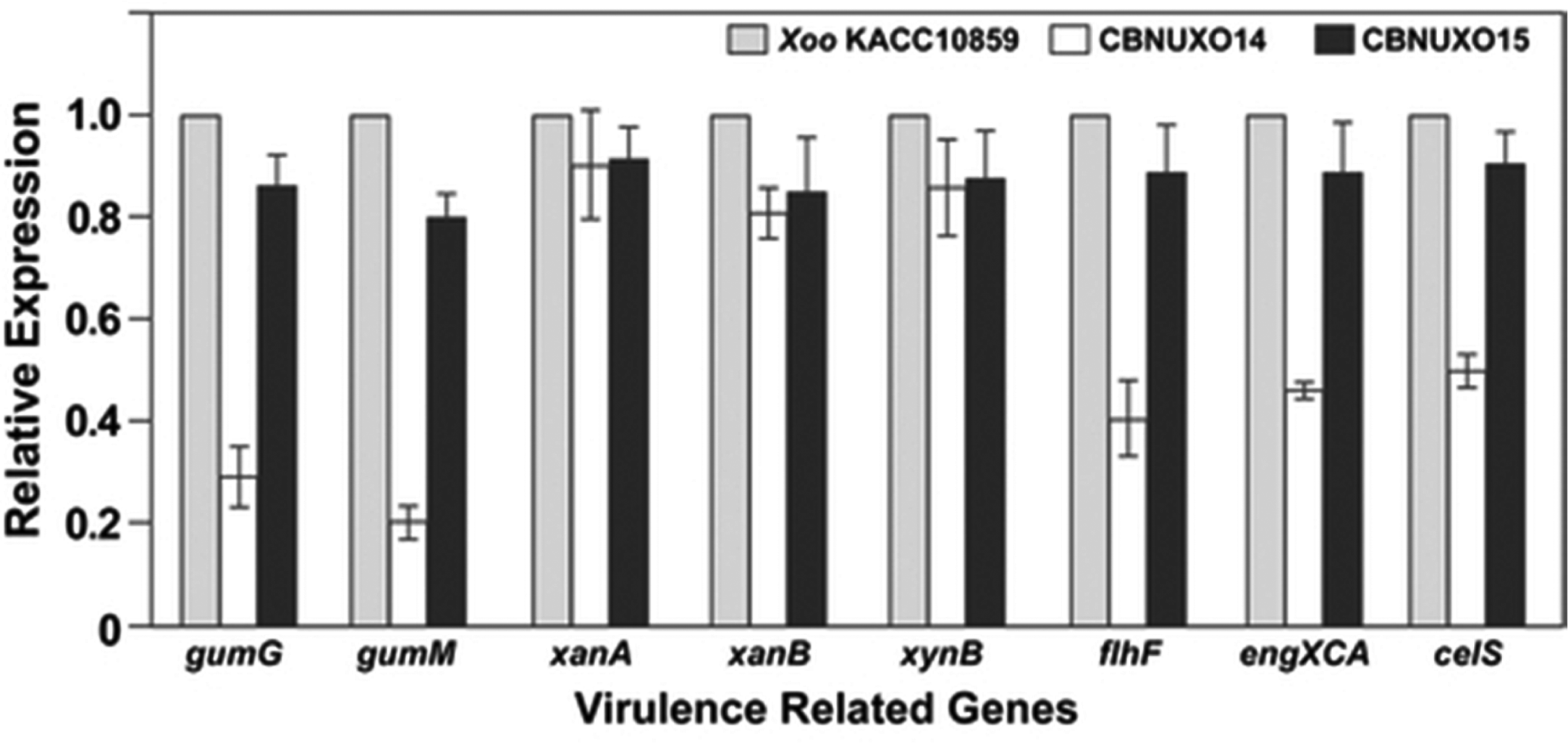
We measured the expression of genes involved in Xoo pathogenicity using quantitative real time-polymerase chain reaction (qRT-PCR) to test if the phenotypes observed in the CBNUXO14 strain resulted from changes in gene expression. The expression of related genes in EPS production (gumG and gumM), swarming motility (flhF), and cellulase activity (engXCA and celS) was strongly reduced in the rpfE mutant strain, CBNUXO14, compared to the WT KACC10859. The expression of LPS-related genes (xanA and xanB) and the xylanase production-related gene (xynB) did not differ (Fig. 4). The complementation strain, CBNUXO15, did not differ from the WT strain KACC10859 in the expression of these genes (Fig. 4). These results show that the knock-out of Xoo rpfE decreased the expression of genes involved in EPS synthesis, motility, and cellulase production, which probably resulted in a decline in Xoo virulence.
rpfE is not involved in DSF synthesis
To test the RpfE requirement for DSF synthesis, we compared DSF production in rpfC, rpfE, and rpfF mutant strains with that in the WT strain (Fig. 5). Using thin layer chromatography (TLC), we analyzed DSF (cis-11-methyl-2-dodecenoic acid) in extracts of suspension medium from each strain culture. The retention factor (Rf) value for the DSF standard was 0.683 (lane 1 in Fig. 5). WT, DSF-overexpressing mutant (rpfC) and rpfE mutant strains produced visible spots under UV illumination that migrated with the DSF standard. The calculated Rf values of the spots from each mutant strain closely matched that of the DSF standard (wild type, 0.675; rpfC, 0.678; rpfE, 0.675), suggesting that rpfC and rpfE mutant strains produced DSF (Fig. 5). The rpfF mutant strain, however, does not produce DSF because rpfF is required for DSF synthesis and the strain is DSF- deficient (He et al., 2010). The result of the TLC analysis indicated that rpfE deficiency does not affect DSF synthesis (Fig. 5).
The rpfE mutant cannot use sucrose and xylose as carbon source
Pathogenic bacteria in the host environment must assess local conditions, including nutrient availability, and adjust their virulence functions accordingly (Feng et al., 2009; Nakamura et al., 2006; Zhao et al., 2012). We tested the growth of the rpfE mutant strain under different carbon source to determine the response of rpfE expression to nutrient conditions. The rpfE mutant strain showed almost no growth on M9 medium containing sucrose and xylose, whereas the growth rate recovered on M9 medium containing glucose (Fig. 6). These results suggested that rpfE expression is sensitive to the carbon source and may regulate genes involved in carbon source utilization, especially for sucrose and xylose.
Discussion
The rpfE gene was reported first in Xcc as a member of the rpf gene cluster (Dow et al., 2000), and genome sequences of xanthomonads and the closely related X. fastidiosa showed that rpfE is a well-conserved gene at this location (Chatterjee et al., 2008; Lee et al., 2006). The biological function of the gene, however, has not been well characterized. In this study, we showed that rpfE is required for the virulence of Xoo by modulating cellulase activity and swarming motility without change of DSF production. The rpfE knock-out mutant also could not utilize sucrose and xylose as carbon source efficiently. These changes by RpfE deficiency most likely decreased virulence ability, especially in the initial stages of infection.
RpfE was reported to control the production of some extracellular enzymes and EPS in Xcc without significant change in virulence; Mutation in rpfE decreases EPS and endoglucanase levels but increases polygalacturonate lyase activity in Xcc (Dow et al., 2000). The endoglucanase of the paper is the same enzymatic activity to cellulase in this paper since the assays for two enzyme activities were identical. In our investigation, however, the rpfE mutation significantly decreased virulence in Xoo. This disparity is probably due to differences in the pathogenic mechanisms of the 2 pathogens. For instance, the Xoo genome sequence includes no orthologs of the pectate lyase or polygalacturonase genes and those enzyme activities were not detected in Xoo (Lu et al., 2011). The absence of the pel and peh genes in Xoo is consistent with the capacity of Xoo to infect only rice plants, which have cell walls containing large amounts of xylan but only low levels of pectic substances (Lu et al., 2011; Yamaguchi et al., 2009).
Phytopathogenic bacteria are confronted by a formidable barrier, the cell wall of the host. Cell wall degrading enzymes are therefore important virulence factors of phytopathogenic bacteria (Jha et al., 2005). Pathogenic bacteria induce expression of genes encoding extracellular enzymes for host cell wall degradation in the first 12 to 72 h after infection (Chalupowicz et al., 2010). The extracellular enzymes including cellulase of phytopathogenic bacteria assist in releasing nutrients from the host plant cell walls (Dow et al., 1990, 1998; Gough, 1988; Hu et al., 2007; Roberts et al., 1988; Walker, 1994). The rice pathogen, Xoo uses a bacterial Type 2 secretion system to secrete several cell-wall-degrading enzymes including 2 cellulases (ClsA and EglXoB), cellobiosidase (CbsA), lipase/esterase (LipA), and xylanase (He et al., 2007; Jha et al., 2007; Rajeshwari et al., 2005; Ray et al., 2000). The sharp decline in cellulase activity with consequent growth retardation in the rpfE mutant strain, not in other rpf- mutants of Xoo (Jeong, 2008), explains the decreased virulence ability of the rpfE mutant. After infection, Xoo spreads through the vascular system of rice, which is the vital conduit for water and nutrients. Considering the evidence that rice uses sucrose as a major from of sugar transport (Elling et al., 1993), rpfE-deficiency hinder sucrose use after infection of the rice vascular system and result in retardation of disease progression in the early stage.
Reduced virulence of pathogenic bacteria has been associated with a lack of EPS as well as cellulase (Hu et al., 2007; Jha et al., 2007; Tang et al., 1996). EPS influences virulence in both Xcc and Xoo. It plays a significant role in the virulence of Xoo (Dharmapuri et al., 2001; Hu et al., 2007), while it is not essential for virulence in Xcc (Dharmapuri and Sonti, 1999). Swarming ability is also an important virulence factor in Xoo (Cho et al., 2011; Shen et al., 2001). The Xoo rpfE mutant showed a reduction in swarming motility in our study (Table 1). Taken together, these results suggest that a deficiency of critical virulence factors, e.g., cellulase, EPS, and motility may explain the reduction in virulence of the Xoo rpfE-mutant strain.
The DSF and Rpf proteins play critical roles in the process of host infection (Chatterjee et al., 2008; He et al., 2010; He et al., 2007; He et al., 2006; Jeong et al., 2008). It has been known that Rpf proteins control the production of pathogenicity factors such as EPS, LPS, and extracellular enzymes (Dow et al., 2006; Dow et al., 2000; Jeong et al., 2008; Slater et al., 2000). The reduction of virulence and decrease in the production of virulence factors in rpfE are consistent with the effects of mutations in other rpf genes. However, our results suggested that Xoo rpfE is not involved in DSF production. This irrelevance between one of the rpf genes and the rpf system was shown in Xcc rpfA encoding aconitase (Wilson et al., 1998) and Xcc rpfE (Dow et al., 2000).




 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






