

The total global agricultural damage caused by plant-parasitic nematodes is estimated to be $100 billion USD per year, among which the most important are root-knot nematodes (RKNs: Meloidogyne spp.; Oka et al., 2000). RKNs are distributed globally, infecting more than 2,000 plant species and reducing global crop yields by about 5%, mainly through root-knot gall formation and nutritional deprivation (Sasser, 1977).
RKNs are commonly controlled by chemicals such as soil fumigant and non-fumigant nematicides; however, these have not been able to achieve full control of RKNs due to the soilborne nature of these nematode pests. In addition, chemical methods are very toxic to humans, animals, and sometimes even plants, and can cause soil and water pollution against which new nematicide production techniques are urgently required (Oka et al., 2000; Osman and Viglierchio, 1981).
Biological control offers a good alternative to chemical control, and provides efficient control with no or little hazard to the soil environment (Noling and Becker, 1994). A variety of microorganisms and natural enemies antagonistic to soil nematodes exist in the soil, including bacteria, fungi, predatory nematodes, and mites. These microbial antagonists and natural enemies are usually found in “nematode-suppressive” soils, in which their damage to plants is below economic threshold levels. Microbes may propagate and spread in the soil for a long time without permeating host plants, and some microbes even enhance plant growth. Therefore, the application of antagonistic soil microbes is expected to effectively control nematodes and act as useful biological control agents against RKNs (Oka et al., 2000).
In Korea, Meloidogyne incognita, M. arenaria, and M. hapla are the major RKNs. M. incognita and M. arenaria are the most common RKN species distributed in warmer regions such as greenhouse soils in Korea (Kim, 2001; Kim et al., 2001b). However, M. hapla favors cooler temperatures and is widely distributed in open-field soils, decreasing qualitative and quantitative production loss of root crops such as ginseng, Codonopsis lanceolata, and carrot in Korea (Kim, 2001; Park et al., 1994, 1995). Control of RKNs under greenhouse conditions has been accomplished using cultural and chemical methods, including treatment with nematicides, admixtures of soil, rotation, and flooding (Kim and Choi, 2001; Kim et al., 2001a). These control measures are more effective against M. incognita and M. arenaria in greenhouse soils than against M. hapla in open-field soils because of the confined cultivation of high-valued crops under controlled environmental conditions in greenhouses (Brown, 1978). Studies on breeding or selection of vegetables resistant to M. incognita and M. arenaria have been conducted, in which several species and lines of vegetables have been used as genetic resources for breeding and controlling RKNs (Kim et al., 2013; Seo et al., 2014).
However, studies on the control of M. hapla have not been conducted using the control measures mentioned above in Korea due to difficulties in their use for crop cultivation under open-field conditions (Brown, 1978). Thus, alternative methods to control M. hapla are required, for which antagonistic soil microbes with high nematicidal activities, rapid multiplication rates, and durable resistance to environmental stresses can be used for sustainable control capacity over a long period under open-field conditions. Therefore, the purpose of this study was to identify soil microbes that can function as biocontrol agents for M. hapla through in vitro selection of antagonistic microorganisms, and to investigate their biocontrol efficiency in vivo against RKNs. This environmentally friendly method to control RKNs may be applicable to current situations in which carrots are being damaged by the pest M. hapla (Seo et al., 2014).
Materials and methods
Preparation of nematode inoculum
RKNs used in these experiments were isolated from root galls of ginseng cultivated in Jinan, Jeon-buk Province, Korea, and identified as Meloidogyne hapla based on analysis of 28S rRNA sequences following the method described by Oh et al. (2009; Kim, unpublished data). Four-week-old tomato plants (Solanum lycopersicum cv. Rutgers) grown in a growth chamber were inoculated with second-stage juveniles (J2) of M. hapla and cultivated at approximately 25±2°C in a greenhouse. Seven weeks following inoculation, whenever inoculums were required, tomato plants were uprooted and root systems were carefully washed with running tap water to remove adhered soil. Egg masses of M. hapla were handpicked with the help of the forceps and were placed on a Baermann funnel for 3-5 days to obtain hatched out J2 (Southey, 1986). The inoculum concentrations of J2 were adjusted to required densities using sterile distilled water (SDW), and were employed for in vitro and pot experiments in our study.
In vitro screening of antagonistic bacteria for nematicidal activity against M. hapla
Soil samples were collected from various fields and mountainous areas, from which bacteria were isolated through the dilution plating method (Dhingra and Sinclair, 1985), resulting in a total of 523 bacterial isolates. These bacterial isolates and 19 Paenibacillus strains solated from the 4-year-old Korean ginseng roots with rot symptoms (Jeon et al., 2003); in total, 542 bacteria were subjected to in vitro nematicidal activity screening.
For the first screening bacterial isolates were cultured in brain heart infusion (BHI) broth (BHIB; Conda S.A., Madrid, Spain) at 28°C for 2 days agitating at 200 rpm in a shaking incubator. The bacterial cultures were diluted with SDW to concentrations of 2% and 1%, and then 100 μL of the diluted bacterial culture was transferred to each well of a 96-well Microtest™ Tissue Culture Plate (Becton Dickinson Labware, Franklin Lakes, NJ, USA) with three replications, into which 50-70 M. hapla J2 were placed. After 24 h of exposure, M. hapla J2 were examined under a low power stereoscopic microscope, and nematodes were considered dead if they were straight, stiffened, and did not move when probed with a fine needle, compared to flexible and responsive (to the fine needle touch) nematodes that were considered alive (Cayrol et al., 1989).
For the second screening, the bacterial isolates selected in the first screening (based on their high nematicidal activities) were screened again for nematicidal activity using the bacterial cell suspensions without culture filtrate (culture filtrate-free bacterial cell suspensions) that may contain nematicidal compounds produced during culture. For this, the bacterial cultures grown in BHIB at 28°C for 48 h in a shaking incubator (at 200 rpm) were centrifuged at 5,162 × g to collect bacterial cells in a pellet by removing supernatant from bacterial cultures. The collected bacterial cells in the pellet were suspended in SDW and diluted to the proper concentrations, for which their colony-forming units (CFU) were examined through the dilution plating method (Dhingra and Sinclair, 1985). Nematicidal activity of the bacterial suspensions was examined using 100 μL of the bacterial suspensions with concentrations of 1×106 CFU/mL and 1×108 CFU/mL using the same methods mentioned in the first screening of bacterial cultures for nematicidal activity.
Identification of antagonistic bacterium selected by in vitro screening
One bacterium (isolate C1-7) out of 542 bacterial isolates was selected in the first and second screening tests as a potential candidate for the biological control of M. hapla. The bacterium C1-7 was identified based on colony morphology, gram staining, carbon source assimilation, and 16S rRNA gene sequencing analysis. Colony morphology was examined by visualizing the colonies formed after 3 days of culturing on BHI agar. Gram staining was conducted following the method previously described by Schaad et al. (2001). Bacterial morphology was examined under an energy-filtering transmission electron microscope (Libra 120, Carl Zeiss, Oberkochen, Germany) operated at an accelerating voltage of 120 kV following negative staining with 2% phosphotungstic acid (PTA). Carbon source assimilation of the bacterial isolate was examined with a Biolog GN test kit (Biolog Inc., Hayward, CA, USA). For 16S rRNA gene sequencing analysis, chromosomal DNA was extracted from the 2-day-old bacterial colony on BHI agar using a FastDNA spin kit (MP Biomedicals, Santa Ana, CA, USA). In addition a polymerase chain reaction (PCR) was performed using universal primers 27mF (5′-GAGTTTGATCMTG GCTCAG-3′) and 1492mR (5′-GGYTACCTTGTTACG ACTT-3′) in a MyCycler Thermal Cycler system (Bio-Rad, Hemel Hempstead, Herts, UK) for the amplification of the 16S rRNA gene (Yoon et al., 1998). The amplified PCR products were purified using a Wizard SV Gel and PCR cleanup system (Promega, Madison, WI, USA), and were then used to analyze for 16S rRNA gene sequences with an ABI PRISM BigDye™ Terminator Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA). Results were compared with ribosomal DNA sequences registered in GenBank using a BLASTN program for bacterial identification (Thompson et al., 1994).
Growth characteristics of C1-7 in different culture media
To determine the optimal population density and culture media for the most efficient application of isolate C1-7, growth curves were examined every 6 h for 114 h after inoculation on four different media including nutrient broth (NB; Conda S.A.), BHIB, Luria-Bertani (LB) broth (LBB; Difco, Detroit, MI, USA), and Bacto™ tryptic soy (TS) broth (TSB; Becton, Dickinson and Company, Le Pont de Claix Cedex, France). For this step, the numbers of CFU were examined by the dilution plating method (Dhingra and Sinclair, 1985).
Nematicidal activity of C1-7 cultured in different media
Four different culture media (NB, BHIB, LBB, and TSB) were used to examine bacterial growths and to test for the nematicidal activity of C1-7. One milliliter of bacterial solution was taken from the bacterial culture at intervals of 6 h after the initial inoculation. Samples were diluted with SDW to various concentrations (0.5%, 1.0%, 2.0%, and 5.0%), and then 100 μL of diluted bacterial solution was transferred to each well of a 96-well Cell Culture Plate (F-Type; SPL Life Sciences, Pocheon-si, Gyeonggi-do, Korea) into which 50-70 J2 of M. hapla were placed. After 24 h of exposure, dead and living M. hapla J2 were counted under a low-power stereoscopic microscope as mentioned above in the in vitro screening.
Effect of the bacterial culture filtrate on nematode mortality
The culture filtrate of isolate C1-7 was also examined for nematicidal activity. The bacterial isolate was grown in BHIB at 28°C in a shaking incubator (200 rpm) for 18 h (just after the end of the exponential growth phase) and 48 h (in stationary phase). The bacterial cultures were centrifuged at 5,162×g to separate the culture supernatant from the bacterial cells condensed in pellet, and the culture supernatant was filtered through a sterilized 0.22-μL Millex Filter Unit syringe (Millipore, Bedford, MA, USA). The culture filtrate was subjected to three heat treatment conditions including a no-heat treatment, heat treatment at 80°C for 30 min, and heat treatment at 121°C for 20 min. Then, each sterilized culture filtrate was diluted with SDW to various concentrations, and 100 μL of the diluted culture filtrate was transferred to each well of a 96-well Cell Culture Plate (F-Type) into which 50-70 M. hapla J2 were placed. After each 24 h and 48 h of exposure, J2 mortality was examined as above.
Effect of the antagonistic bacterium on the control of M. hapla in pot experiments
C1-7 was tested for biological control of M. hapla on host plants, carrot (Daucus carota subsp. sativus) cv. Shinheukjeon-Ochon and tomato, both of which are highly susceptible to the RKNs (Bélair, 1984; Hartman and Sasser, 1985). The carrot is a representative host crop of M. hapla, and control is urgently required for crop cultivation. The tomato cv. Rutgers is also susceptible to M. hapla and other Meloidogyne species (Hartman and Sasser, 1985), and was used for comparison in our study. Four-week-old seedlings were planted in 8-cm-diameter plastic pots that were filled with a sterilized sand-bed soil (1:1) mixture, and each plant was inoculated with M. hapla by dispending 5 mL of the nematode suspension containing 200 J2/mL per pot. For the biological control of the nematode, 10 mL of C1-7 culture grown in BHIB at 28°C in a shaking incubator (200 rpm) for 3 days and diluted with SDW to the concentration of 1×108 CFU/mL was poured around the plant rhizosphere at the same time as nematode inoculation. Plants with neither nematode inoculation nor bacterial treatment served as controls. Fresh BHIB (10 mL; diluted as much as the bacterial culture) was poured around the plant rhizosphere to examine the effect of the medium on nematode control. Each treatment was replicated five times and the pot experiments were conducted at 25±2°C under greenhouse conditions. Plants were watered sufficiently every other day. Four weeks after treatments, plants were carefully uprooted, and root systems were gently washed with running tap water to remove adhered soil. The severity of root galling on plants infected with M. hapla was assessed on a 0-5 rating scale according to the percentage of galled tissues, in which a rating of 0 = 0-10% galled roots; 1 = 11-20%, 2 = 21-50%, 3 = 51-80%, 4 = 81-90%, and 5 = 91-100% (Barker, 1985). Egg masses of M. hapla were handpicked from infected roots using forceps, and an egg mass index was assigned to each count using a rating scale of 0 = no egg mass, 1 = 1-3 egg masses, 2 = 4-10 egg masses, 3 = 11-30 egg masses, 4 = 31-100 egg masses, 5 = >100 egg masses per root system (Roberts et al., 1990).
Effect of the antagonistic bacterium on histopathological changes in plants infected with M. hapla
Root segments with various degrees of galling were cut off from carrot and tomato roots inoculated with M. hapla, both with and without the bacterial treatment. The root segments were fixed in Karnovsky's fixative consisting of 2% glutaraldehyde and 2% paraformaldehyde in 0.05 M cacodylate buffer (pH 7.2) for 48 h at 4°C (Karnovsky, 1965). The fixed specimens were washed in 0.05 M cacodylate buffer (pH 7.2) at 4°C three times each for 10 min. The specimens were then post-fixed in 1% osmium tetroxide and 0.05 M cacodylate buffer for 1.5 h at 4°C before brief rinsing with distilled water. After en bloc staining in 0.5% uranyl acetate for 30 min at 4°C, the specimens were dehydrated in an ethanol series of 30%, 50%, 70%, 80%, and 90% each for 10 min, and finally three times in 100% ethanol for 10 min each. For further dehydration, the specimens were treated with propylene oxide at room temperature two times each for 15 min. The specimens were infiltrated with a 2:1 mixture of propylene oxide and Spurr's epoxy resin for 4 h, then a 1:1 mixture overnight, followed by 100% Spurr's epoxy resin under a vacuum for 4 h in a desiccator. The specimens were embedded in fresh Spurr’s epoxy resin in a dental mold, which was placed at 70°C for 8 h in a dry-oven for polymerization. The specimens embedded in the resin were cut to slices of 1,000 nm thickness with a glass knife on an ultramicrotome (MT-X, RMC, Tucson, AZ, USA). The sections were stained using decuple diluted toluidine blue working solution consisting of 1% toluidine blue in 2% sodium borax, and observed under a compound light microscope (Axiophot, Carl Zeiss, Oberkochen, Germany).
Results
In vitro screening of antagonistic bacteria for nematicidal activity against M. hapla
Under in vitro conditions, the bacterial cultures of a total of 542 bacterial isolates grown in BHIB for 48 h were diluted to 1.0% and 2.0%, and screened for nematicidal activity against M. hapla. Among these bacterial cultures, 23 bacterial isolates had a high nematicidal activity, providing near to 100% mortality at 2.0% concentration (except for two isolates: S-2 and S-4) and was still effective at 1.0% bacterial isolate (except for three isolates: S-2, S-4, and S-7; Fig. 1A). In the second in vitro screening experiment with culture filtrate-free bacterial cell suspensions at concentrations of 1×106 CFU/mL and 1×108 CFU/mL, all isolates except for C4-19 and S-5 showed 100% nematode mortality at a concentration of 1×108 CFU/mL; however, only three isolates (C1-7, S-2, and S-4) provided 100% mortality at 1×106 CFU/mL (Fig. 1B), indicating that isolate C1-7 had the highest nematicidal activity of 100% morality at the lowest concentrations of both the bacterial culture and suspension .
Identification of the antagonistic bacterium
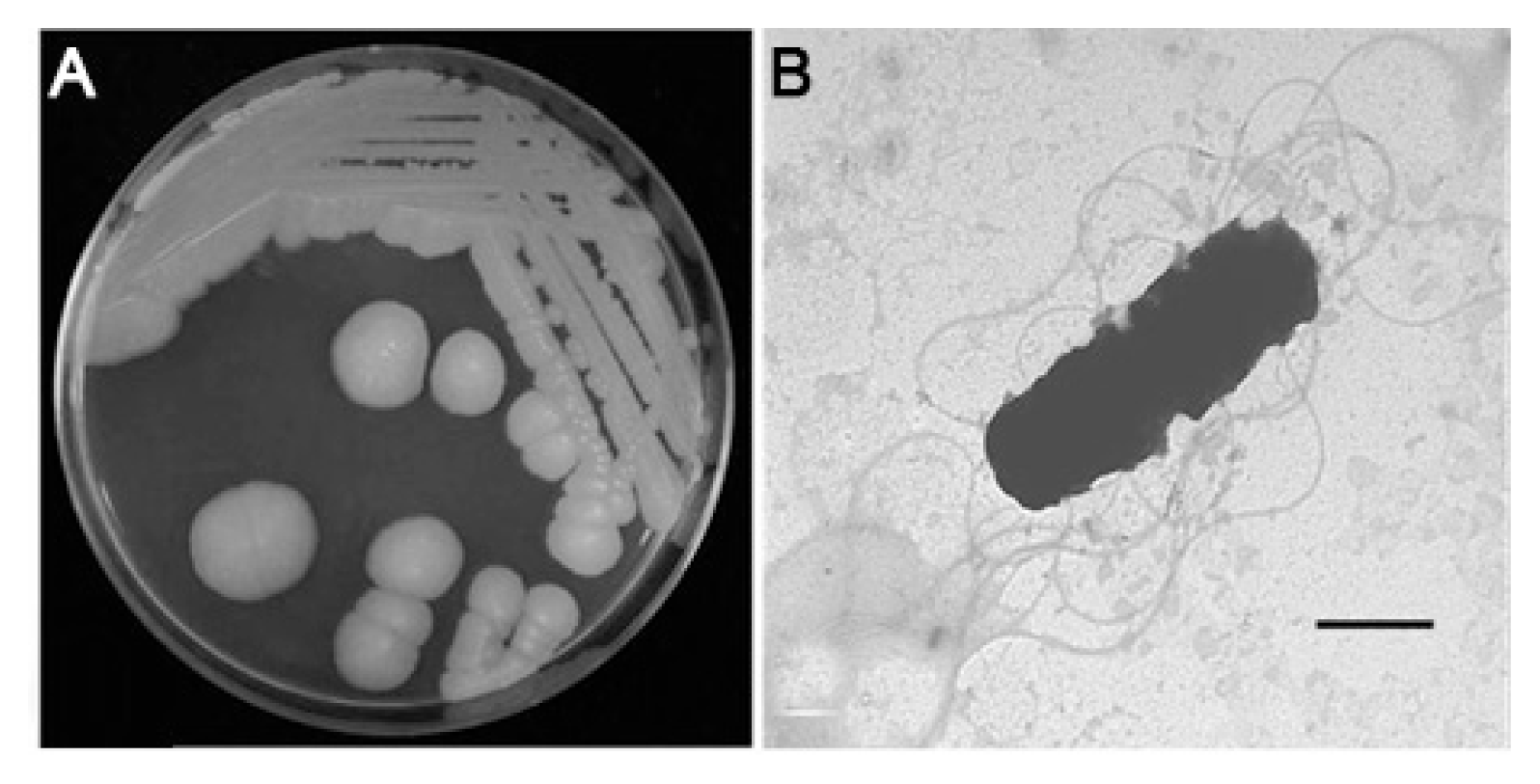
C1-7 formed large circular, cream-colored, flat colonies with an undulate margin after 3 days culture on BHI agar (Fig. 2A), and it was gram-positive, rod-shaped, and had more than four peritrichous flagella (Fig. 2B), showing the most similarities with the genus Bacillus (Schaad et al., 2001). The carbon source assimilation of the bacterial isolate examined using a Biolog assay revealed the utilization of 15 carbon sources including glycerol, but not 34 other carbon sources including mannitol, revealing the highest similarity (92.6%) with Bacillus cereus (data not shown). The analysis of 16S rRNA gene sequences for C1-7 revealed a maximum similarity (100%) to those of B. cereus strains listed in GenBank with accession numbers KC113613.1, JX848326.1, JX994144.1, and JX994107.1 (data not shown), including an E-value of 0.0, confirming the bacterium’s identity as B. cereus.
Growth characteristics of the antagonistic bacterium on different culture media
For all media examined, the lag phase observed in population growth was hardly noted in the culture of the bacterial isolate C1-7; however, its population increased rapidly up to 18 h after inoculation in all media examined, revealing an exponential phase with populations reaching concentrations of 1×108 CFU/mL ~1×109 CFU/mL (Fig. 3). Population growth rates were differentiated after the exponential phase depending on the media, revealing a stationary phase from 36 h through 60 h after inoculation (or to the end of culturing with no death phase) in all but NB, maintaining a bacterial density of approximately 0.5×109 CFU/mL in BHIB and approximately 1.0×108 CFU/mL in LBB and TSB. In NB, the bacterial population decreased significantly after the exponential growth phase, with no noticeable duration of the stationary phase, to a final population of less than 1.0×106 CFU/mL.
Nematicidal activity of the antagonistic bacterium cultured in different media
No nematicidal activity was observed for the bacterial cultures at all concentrations of NB and at the concentration of 0.5% of all culture media used in this study (Table 1). Only 5.0% of the bacterial cultures in LBB and 2.0% and 5.0% in TSB showed sufficiently high nematicidal activity throughout the culturing period, while all concentrations except for 0.5% in BHIB showed high nematicidal activity throughout the culturing period, indicating that BHIB is the optimal medium for the growth of C1-7 for the nematicidal activity.
Nematicidal activity of the bacterial culture filtrate with different culture times, heating temperatures, and treatment times
The bacterial culture filtrate displayed much higher nematicidal activity against M. hapla than either BHI or the control (SDW), regardless of culture time, heating temperature, and time exposed to heating (Table 2). The culture filtrate of the bacterial isolate using an 18-h culture time showed no significant difference in nematicidal activity against M. hapla J2 between the exposure times of 24 h and 48 h, regardless of heating temperatures, displaying nearly total nematode mortality at concentrations of 2% and greater. However, the nematicidal activity of the bacterial culture filtrate using a 48-h culture time was higher using an exposure time of 48 h compared to one of 24 h. Total nematode mortality was observed at a concentration of 1% using a treatment time of 48 h, but not with a treatment time of 24 h, for both of which no significant differences in nematicidal activity were detected among the heating temperatures (Table 2).
Effect of the antagonistic bacterium on the control of M. hapla in pot experiments
In the pot experiments under greenhouse conditions, root-knot galls and egg masses formed on carrot and tomato plants inoculated with M. hapla were significantly suppressed by treatment with the C1-7 bacterial culture (1×108 CFU/mL), illustrating that the size of root-knot galls and the number of root-knot galls and egg masses significantly decreased in the C1-7 treatment compared to the nontreatment control (Table 3, Fig. 4). For root-knot gall formation, complete suppression happened in tomato, while for egg mass formation, complete suppression occurred only in carrot; however, 90% suppression of root galling was observed in carrot and 75% suppression of egg mass formation was observed in tomato (Table 3). BHIB treatment also reduced root galling and egg mass formation, especially significantly egg mass formation in tomato; however, its control efficacy was far below that of the bacterial culture.
Effect of antagonistic bacterial treatment on histopathological changes in plants infected with M. hapla
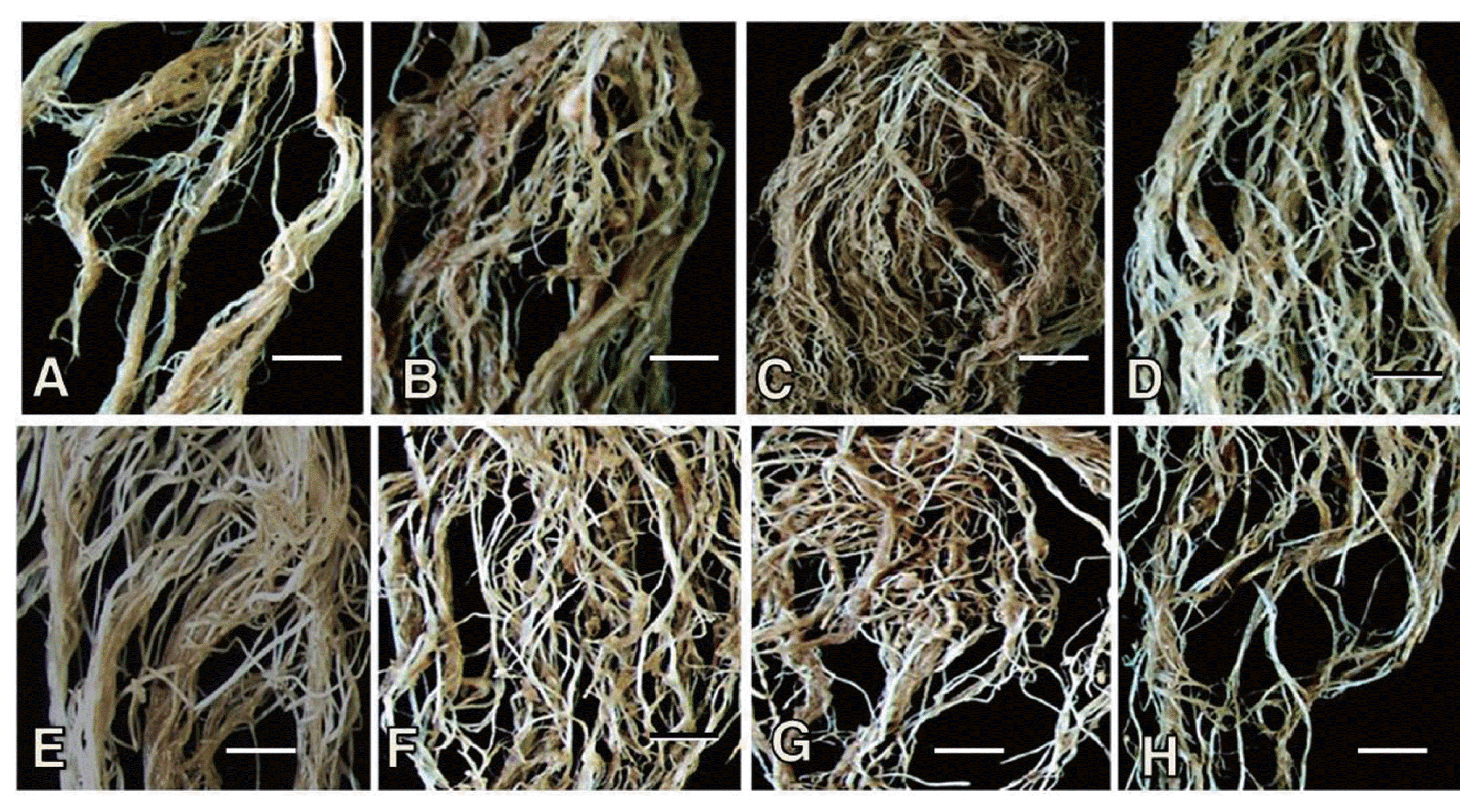
In the control plants with no bacterial treatment, numerous large giant cells were formed in carrot and tomato roots infected with M. hapla, which is characterized by cellular hypertrophy with the formation of multiple nuclei and sometimes cell wall ingrowths (Fig. 5). The proliferation of gall cells was noted around the giant cells formed in the roots with no treatment control. Giant cells were also formed in the root tissues infected with the nematode and treated with BHIB and the bacterial culture; however, they were small in size with no cell wall ingrowths and often appeared degenerative with vacuolar cytoplasm and electron-dense materials (presumed to be chromatin materials) scattered in the giant cells (Fig. 5C, D, G). No giant cell formation was found in the nematode-infected tomato root tissues treated with the bacterial culture (Fig. 5H). Gall cells proliferated in BHIB to the same extent as the untreated control, but did not proliferate with the bacterial treatment, illustrating that giant cell formation was confined in the stele (Fig. 5D).
Discussion
M. hapla is one of the major RKNs in Korea, which cause qualitative and quantitative damage in the production of field crops and root vegetables such as ginseng and carrot (Kim, 2001). Numerous microbes such as Bacillus spp., Paecilomyces lilacinus, and Verticillium chlamydosporium have been tested for the control of M. hapla, since they suppress root galling, egg mass formation, egg hatching, and the survival of J2 individuals (Bonants et al., 1995; Chen et al., 2000; De Leij et al., 1993; Liu et al., 2008; Mennan et al., 2006; Townshend et al., 1989; Viaene and Abawi, 2000).
In this study, C1-7 (identified as B. cereus) showed the strongest nematicidal activity in vitro against M. hapla among the 523 bacterial isolates tested. Complete nematode mortality was exerted by the bacterial culture and culture filtrate-free bacterial cell suspension at lower concentrations, at which concentration most bacterial isolates (excluding C1-7 and two other isolates) showed no nematicidal activities. This indicates that C1-7 may have exerted nematicidal activity at early treatment times, which would be supported by the rapid bacterial growth and durable nematicidal activity observed during the culture in BHIB.
Furthermore, the culture filtrate of B. cereus C1-7 exerted strong nematicidal activity, suggesting that its mode of action for the biological control of the nematode relies on antibiosis (antagonistic activity of the nematicidal materials produced in the bacterial culture), which is similar to other Bacillus species secreting toxoproteins (Carneiro et al., 1998; Zhang et al., 2012). Thus, B. cereus C1-7 may exert nematode control activity immediately upon application due to the nematicidal compounds produced in the bacterial culture, which is supported by the rapid bacterial growth. This may enhance the overall biological control efficacy, which could provide novel biological control agents (Whipps and Davies, 2000).
Furthermore, unlike the toxoproteins that are usually denatured and deprived of their toxicity by heat shock (Leyns, 1995), the bacterial culture filtrate in our study was heat-stable, similar to non-proteinous thermostable toxic compounds produced by some Bacillus spp. during the stationary phase of bacterial growth (Yu et al., 2012). The population density and the nematicidal activity of B. cereus C1-7 remained high for a long period of time during growth in culture medium. These biological and cultural characteristics of the bacterium may provide some advantages for its formulation and practical application in the biocontrol of RKNs under field conditions owing to the stability and perpetuation of the high nematicidal activity.
In pot experiments, the biocontrol efficacy of B. cereus C1-7 was significantly higher in both carrot and tomato plants. However, the specific biocontrol activity against the RKN differed depending on the host plants, demonstrating that the inhibition rates of gall and egg mass formation were higher in tomato and carrot, respectively. This suggests that the biocontrol of M. hapla with B. cereus C1-7 reduces nematode damage and nematode population growth in tomato and carrot, respectively.
Differences in specific biocontrol activity of B. cereus C1-7 was also supported by light microscopy of the root tissues infected with the RKN, showing no or few giant cells and gall cells in the tomato and no or few nematodes in the carrot treated with the bacterial culture. This observation agreed with anatomical aspects of tomato plants infected with M. incognita, when numerous giant cells formed in large gall tissues with gall cell proliferation, and small giant cells formed in the stele of root tissues without noticeable gall cell proliferation (Kim and Ohh, 1990). Variability of the specific biocontrol activity may be associated with biological characteristics of host plants governing responses to nematode infection, as observed in chili pepper plants infected with M. incognita (Moon et al., 2010) and various host and non-host plants infected with Heterodera glycines (Kim et al., 1986; Kim, 1990). Despite the differences in specific biocontrol activity, these histopathological changes indicate that B. cereus C1-7 has a residual or systemic effect against RKNs infecting root tissues that may have been weakened or escaped from nematicidal activity outside of the root, further increasing the biocontrol efficacy and decreasing the crop yield loss by inhibiting root gall formation (Sasser, 1977).
In this study, the modes of the nematicidal activity of B. cereus C1-7 was not examined in detail; however, one can hypothesize that the reductions in root galling and egg mass formation by M. hapla may be attributable to the direct effects of secondary metabolites that injured the nematode juveniles, or the indirect effects that enhanced host plant defense mechanisms to prevent the nematodes from infection, as found in the plant growth-promoting rhizobacteria Paenibacillus strains (Son et al., 2009). B. cereus is gram-positive, rod-shaped, and genetically close to Bacillus thuringiensis, which is commercially used as biological insecticide. Generally, strains of B. cereus are known to be saprophytic and even beneficial as probiotics, representing a promising biological control agent against plant-pathogenic fungi and nematodes (Chang et al., 2007; Handelsman et al., 1990; Huang et al., 2005; Oka et al., 1993; Ryan, 2004; Silva et al., 2004). Some B. cereus strains are harmful to humans and animals, but they are generally known to be self-limiting and noncontagious.
A plant health-promoting rhizobacterium, B. cereus S18, was tested for the control of M. incognita, M. javanica, and M. arenaria on tomato and was able to reduce root galls and egg masses along with significant increases in plant growth, including shoot height and weight, when compared to the untreated control (Mahdy et al., 2000). Therefore, B. cereus C1-7 is thought to represent a promising biological control agent against problems caused by M. hapla in field crop cultivation.



 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






