

Rice is the staple food over half of the world population and its production was estimated approximately 745 million tonnes in 2013 worldwide. Asia ranks highest in rice production but 24-41% of losses occur annually due to the major limiting factor, diseases (Anon., 2014; Savary et al., 2000). More than 70 diseases caused by fungi, bacteria, viruses, and nematodes have been recorded in rice and especially rice blast, bacterial leaf blight, grain rot, and bakanae are the most serious seed-borne diseases reducing yields (Ou, 1985). The bacterial blight caused by Xanthomonas oryzae pv. oryzae enormously affects yield losses 10-50% of total rice production, depending on the situation (Mew, 1992). Other bacterial diseases including seedling blight and rot, leaf sheath rot, leaf browning, panicle blight, bacterial wilt, and grain rot caused by Burkholderia glumae reduced the yield by up to 75% in some cases (Kim et al., 2010; Ura et al., 2006). The yield losses due to bakanae caused by Fusarium fujikuroi were estimated to be approximately 10-50% in the Asian rice-growing areas (Bonman, 1992). For controlling the diseases, chemical fungicides have been widely used in most Asian countries during past decades, but the efficacy of fungicides has decreased recently due to the occurrence of resistance (Yang et al., 2012). Additionally, the misuse of chemical fungicides has adverse effects on the agricultural environment and farmers, so the development of alternative control measures, specifically, biological controls using antagonistic microorganisms, has been attempted in many Asian countries (Gnanamanickam, 2009).
Biological control using antagonistic bacteria would be environmentally sound and can be implemented as an integrated disease management tool. Various bacterial genera, such as Bacillus, Burkholderia, Lysobacter, Pantoea, Pseudomonas, and Streptomyces have been used as biocontrol agents for controlling diseases of many crops, but a few studies have been reported on the biological control of rice diseases (Bouizgarne, 2013; McSpadden Gardener, 2010). The multiple use of antagonistic bacteria Streptomyces and Bacillus species has been attempted for the control of rice sheath blight (Sung and Chung, 1997). Rice blast and sheath blight were controlled effectively by Bacillus vallismortis EXTN-1 and two antagonistic bacterial strains, Pseudomonas fluorescens mc75 and pc78 (Choi et al., 2006; Park et al., 2006). Bakanae and seedling rot of rice caused by Fusarium moniliforme and F. fujikuroi were also controlled by antagonistic P. fluorescens and Bacillus cereus (Kazempour and Elahinia, 2007; Rosales and Mew, 1997). Among the diverse antagonistic bacteria, several Bacillus species have been developed as commercial bio-pesticides, because Bacillus species can produce endospores and persist successfully in natural environments for a long period after treatment (Hu et al., 2011).
Bacillus species widely used for biological control of many plant diseases in different hosts include Bacillus amyloliquefaciens, Bacillus subtilis, Bacillus pasteurii, B. cereus, Bacillus pumilus, Bacillus mycoides, and Bacillus sphaericus (Kloepper et al., 2004; McSpadeen Gardener, 2010). B. subtilis GB03 and B. amyloliquefaciens IN937 were demonstrated to control the bacterial pathogen, Erwinia carotovora subsp. carotovora in Arabidopsis (Ryu et al., 2004). B. cereus AR156 and B. subtilis were also demonstrated to control Pseudomonas syringae pv. tomato DC3000 successfully in Arabidopsis by inducing resistance (Niu et al., 2011). Some of these Bacillus species have been well characterized in terms of their anti-fungal, anti-bacterial, plant growth-promoting, and resistance-inducing activities in host plants (Park et al., 2009; Ryu, 2013). Diverse species of Bacillus have been isolated from various terrestrial and halophytic plants and some of them have been shown to be endophytic (Bibi et al., 2011, 2012). The genus Bacillus currently contains 299 species and more than 30 species have been reported to be novel during the last 5 years, based on polyphasic studies, including 16S rRNA gene sequences, DNA-DNA hybridization analyses, fatty acid profiles, and physical and biochemical tests (Parte, 2014).
In this study, two Bacillus strains YC7007 and YC7010T isolated from the roots of rice were examined in developing a novel biocontrol agent with multifunctional activities and further characterized for identification by a polyphasic approach. The control efficacy of the novel strain YC7007 was also investigated through induction of resistance against bacterial blight and panicle blight as well as growth promotion in rice.
Materials and Methods
Isolation and cultivation of endophytic bacteria
Endophytic bacterial strains were isolated from the rice roots collected at the paddy field of Gyeongsang National University Farm, Jinju, Korea. For the isolation, sample pieces were washed several times with running tap water and surface-sterilized with 70% ethanol for 5 min, 1.2% NaOCl solution for 10 min, and finally washed with sterile distilled water several times. To confirm the successful disinfection, washed segments were placed on one-tenth strength tryptic soy broth agar (1/10 TSA) at 28┬░C for 3 days and bacterial growth was observed. After confirmation of no bacterial colonies, the sample segments were again sterilized by 70% ethanol for a few seconds and ground in autoclaved distilled water with a sterile mortar and pestle (Bibi et al., 2012). Aliquots were 10-fold serially diluted in autoclaved distilled water and the diluted samples were kept in a hot water bath (80┬░C) for 10 min to select endospore-forming bacterial species. An aliquot (0.1 ml) was plated on 1/10 TSA media supplemented with cycloheximide (50 ╬╝g/ml) and the plates were incubated at 30┬░C for 3 days. The bacterial colonies grown on the medium were selected on the basis of distinct colony morphology. The bacterial strains isolated purely were subcultured on 1/10 TSA media and stored at ŌłÆ70┬░C for further use. For cultivation of bacterial strains, medium containing 10 g protease peptone, 10 g yeast extract, 4 g NH4Cl, 4 g MgSO4, 10 g glucose, and 15 g agar per liter distilled water was prepared.
Antagonistic activity against fungal and bacterial pathogens
The endophytic bacteria were tested against important plant fungal pathogens, such as Alternaria panax KACC 42461, F. fujikuroi KACC 44022, F. oxysporum KCTC 16909, Sclerotinia sclerotiorum GSCC 50501, Pythium ultimum GSCC 50651, Bipolaris oryzae KACC 40853, Botrytis cinerea KCTC 6973, Magnaporthe grisea KACC 40415, Botryosphaeria dothidea GSCC 50201, and Rhizoctonia solani KCTC 40101. The antagonistic activity of bacterial strains was determined by measuring the inhibition zone of mycelial growth of the fungal pathogens on potato dextrose agar (PDA) medium using in vitro confrontation bioassay (Bibi et al., 2012). For the antibacterial test, B. glumae KACC10359 and X. oryzae pv. oryzae KACC 10208 were grown on one-half strength R2A and YGC media (10 g glucose, 30 g CaCO3, 5 g yeast extract, and 15 g agar per liter distilled water), respectively and the inhibition activity was tested using a diffusion paper disc method. Culture filtrate of the bacterial strain YC7007 was prepared from the culture broth with different cultivation times by centrifugation (5,000 g, 10 min) and filtration through a Millipore filter (0.2 ╬╝m).
Bioassay of induced resistance against bacterial pathogens
Induction of resistance by strain YC7007 was tested against bacterial blight and panicle blight caused by X. oryzae pv. oryzae KACC 10208 and B. glumae KACC 10359, respectively, in pot tests. Seeds of rice (Oryza sativa L. cultivar Dongjin) were surface-sterilized with 1.2% NaOCl solution for 5 min and 70% ethanol for 5 min, then rinsed three times with sterile distilled water. The seeds were then kept at 30┬░C for 3 days in the dark for germination and the water was changed every day. The germinated seeds were sown in commercial nursery soil (Dasuran Sangto, Youngnong Sun Up, Korea) and placed in a greenhouse for cultivation. Then, 2-week-old seedlings were transplanted into plastic pots (9.5├Ś8├Ś7 cm3) containing about 150 g commercial nursery soils that had been autoclaved for 20 min at 121┬░C twice on two consecutive days. For the preparation of a bacterial suspension of YC7007, bacterial cells were cultivated for two days in the liquid mass culture medium on a rotary shaker (160 rpm, 28┬░C) and harvested by centrifugation (5,000 g, 10 min) and adjusted to different concentrations (5.6├Ś105, 3.6├Ś106, 2├Ś107 cfu/ml) in buffer solution (10 mM MgSO4). The cell suspension (15 ml) of YC7007 was drenched into the plastic pot containing autoclaved soils (150 g) during transplanting 2-week-old rice seedlings at the same time. The optimal concentration of YC7007 (2├Ś107 cfu/ml) was used for testing further resistance induction against bacterial blight and panicle blight. The culture filtrate of strain YC7007 prepared from a 60 h culture broth that was diluted 10 times with the 10 mM MgSO4 and sprayed to the leaves until to droplet for checking its control efficacy. Then, five days after treatment with YC7007, the bacterial pathogens were inoculated. The buffer solution was used as a control. All experiments were conducted using 10 plants with three replicates for each treatment.
Inoculum preparation of bacterial pathogen
The inocula of two bacterial pathogens, B. glumae KACC 10359 and X. oryzae pv. oryzae KACC 10208 were prepared using the cell suspension after cultivation in R2A and YGC media, respectively, at 28┬░C for 24 h on a rotary shaker (160 rpm). The culture broth of B. glumae was centrifuged (5,000 g, 10 min) and cell pellets were suspended in buffer solution (10 mM MgSO4) adjusted to 6├Ś107 cfu/ml. A pin-prick bioassay was conducted for inoculation of the suspension of B. glumae. A bundle of 3-4 pins/needles was dipped into the suspension and then leaves were teased with the needles of bundle. Disease severity was evaluated 5 days after inoculation of the pathogen to the 5-week-old rice seedlings using a 0-3 disease rating scales: 0 = symptomless, 1 = slight infection, few small lesions, 2 = coalesced lesions to browning all over, and 3 = no description available (Cottyn et al., 1996). In case of bacterial blight, the suspension of X. oryzae pv. oryzae was prepared following the same procedure of B. glumae and concentration was adjusted to 1.2├Ś107 cfu/ml. A clipping bioassay for bacterial blight was conducted using scissors that were dipped into the suspension and then used to cut the upper leaves. Disease severity was determined 7 days after inoculation according to a 1-9 scale (Misra et al., 1994). Disease reduction was calculated following the formula: [(disease severity of the control - disease severity of a treatment) / disease severity of the control] ├Ś 100%.
Measurement of growth promotion
Growth promotion of rice by treatment with a bacterial suspension of strain YC7007 (2├Ś107 cfu/ml) was examined during cultivation periods. Bacterial suspension (1 ml) of YC7007 was drenched on the rhizosphere of 5-day-old rice seedlings in test tubes (18 cm length) containing 10 ml 1/2 MS media with 0.8% agar. Growth data were recorded after 7 days of the bacterial treatment (12 days old). In case of the pot tests, 2-week-old seedlings were transplanted into plastic pots containing about 150 g autoclaved nursery soil and the bacterial suspension (15 ml) was drenched at 7 days from transplanting. Growth data were recorded after 9 days of the bacterial treatment (30-day-old plants). Again, the bacterial suspension (15 ml) was drenched at the tillering stage (30 days old) and growth data were recorded at the booting stage after 40 days of bacterial treatment from the tillering stage (70 days old). Ten plants with three replicates for each treatment were conducted for growth promotion activities for the test tubes and pot tests. The buffer solution (10 mM MgSO4) was used for the control treatment.
Phylogenetic analysis based on 16S rRNA gene sequences and DNA-DNA hybridization
The 16S rRNA gene was amplified from the genomic DNA extracted with a commercial extraction kit (Intron Biotech, Seoul, Korea) using bacterial universal primers 27F and 1492R and the purified PCR product was sequenced by GenoTech Inc. (Daejeon, Korea) (Lane, 1991). To ascertain the phylogenetic position of the novel endophytic bacteria and other closely related type strains, the 16S rRNA gene sequences of the strains were compared with sequences obtained from NCBI and the EzTaxon-e database server (Kim et al., 2012). Multiple alignments of the sequences were performed using the CLUSTAL_X software (Thompson et al., 1997). Gaps were edited using the BioEdit program (Hall, 1999). The neighbor-joining method (Saitou and Nei, 1987), maximum-parsimony (Fitch, 1972), and maximum-likelihood algorithms in the MEGA 5.10 software (Tamura et al., 2011) were used to construct phylogenetic trees with bootstrap values based on 1,000 replications (Felsenstein, 1985). To determine the DNA-DNA relatedness values among strains, DNA-DNA hybridization was carried out using the DIG DNA labeling and detection kit (Roche Applied Science) according to the instructions of the manufacturer and the method described previously (Lee et al., 2011).
Morphological, phenotypic and chemotaxonomical characteristics
Cell morphology was observed under a light microscope (Nikon) at 1000├Ś magnification and the presence of a flagellum was investigated by transmission electron microscopy (Hitachi, model H-600) using a cell broth grown at 28┬░C for 24 h in R2A medium. Gram reaction was determined using the bioM├®rieux Gram stain kit according to the manufacturerŌĆÖs instruction. The hydrolysis tests of strains were performed by a standard protocol, using casein, esculin, gelatin, starch, L-tyrosine, urea, Tween 20, and Tween 80 (Reichenbach, 1992; Smibert and Krieg, 1994). Enzyme activities, acid production from different carbohydrates, assimilation of various substrates and growth on carbohydrates were determined using commercial systems-API ZYM, API 20E, API 20NE, and API 50CH kits, respectively, at 28┬░C according to the manufacturerŌĆÖs instructions (BioMeri├®ux). Growth at different temperatures and various pH values (pH 4.0-14.0 at intervals of 0.5 pH units) was investigated after 5 days incubation on R2A broth using appropriate buffers (Xu et al., 2005). Salt tolerance was tested in R2A broth supplemented with 1-14% (w/v, at 1% intervals) NaCl after 5 days incubation at 28┬░C. Duplicate antibiotic-sensitivity tests were performed using filter-paper disc diffusion assays with different antibiotics: 10 ╬╝g ampicillin, 30 ╬╝g chloramphenicol, 10 ╬╝g penicillin, 10 ╬╝g gentamycin, 30 ╬╝g kanamycin, 30 ╬╝g vancomycin, 30 ╬╝g streptomycin, and 30 ╬╝g tetracycline (Yasir et al., 2010). Preparation of cell walls and analyses of peptidoglycans were carried out using methods described by Schleifer (1985) with the modification that TLC was performed on cellulose sheets rather than by paper chromatography. For the analysis of cellular fatty acids, bacterial strains were cultivated in R2A broth at 28┬░C and the microbial cells were harvested in the mid-exponential growth phase (OD600 = 0.4-0.5). The analysis of fatty acid methyl esters was performed according to the instruction of the microbial identification system (MIDI; Microbial ID, Inc.). Extracts were analyzed by GC (Agilent 6890) and identified by comparing the fatty acid profiles with the TSBA 40 database provided with the Sherlock software (ver. 4.0). Amino acids of whole-cell hydrolysates of bacterial strains were extracted and analyzed according to the method of Staneck and Roberts (1974). Isoprenoid quinones were extracted and analyzed using reverse-phase HPLC according to the method described by Komagata and Suzuki (1987). For the measurement of G+C content of the chromosomal DNA, the genomic DNA of strain YC7010T was extracted and purified as described (Ausubel et al., 1995). It was then enzymatically degraded into nucleosides and G+C content was determined with a reverse-phase C18 column (Mesbah et al., 1989). Polar lipids were extracted by the modified method of Minnikin et al. (1984) and separated by TLC on Merck Kieselgel 60-HPTLC. Aminolipids were detected by spraying the plate with 0.2% (w/v) solution of ninhydrin in butanol saturated with water followed by heating at 105┬░C for 10 min (Ross et al., 1985). Phospholipids were detected by spraying the plate with Zinzadze reagent of Dittmer and Lester (1964). Glycolipids were detected with 1-naphthol spray reagent by heating at 100┬░C for 3-5 min (Jacin and Mishkin, 1965). The presence of phosphatidylcholine was detected with Dragendorff reagent (Sigma-Aldrich; St. Louis, Mo). Total lipid profiles were detected by spraying with phosphomolybdic acid solution (Sigma-Aldrich; St. Louis, Mo) followed by heating at 150┬░C for 10 min.
Statistical analysis
Data were analyzed by analysis of variance designed with a complete randomized single factor for in vitro, complete randomized block design for in vivo, and mean differences were compared using DuncanŌĆÖs multiple range test (DMRT). The SPSS software (ver. 17; SPSS Inc. in Chicago) was used for all analyses.
Results
Antagonistic activity of isolated strains
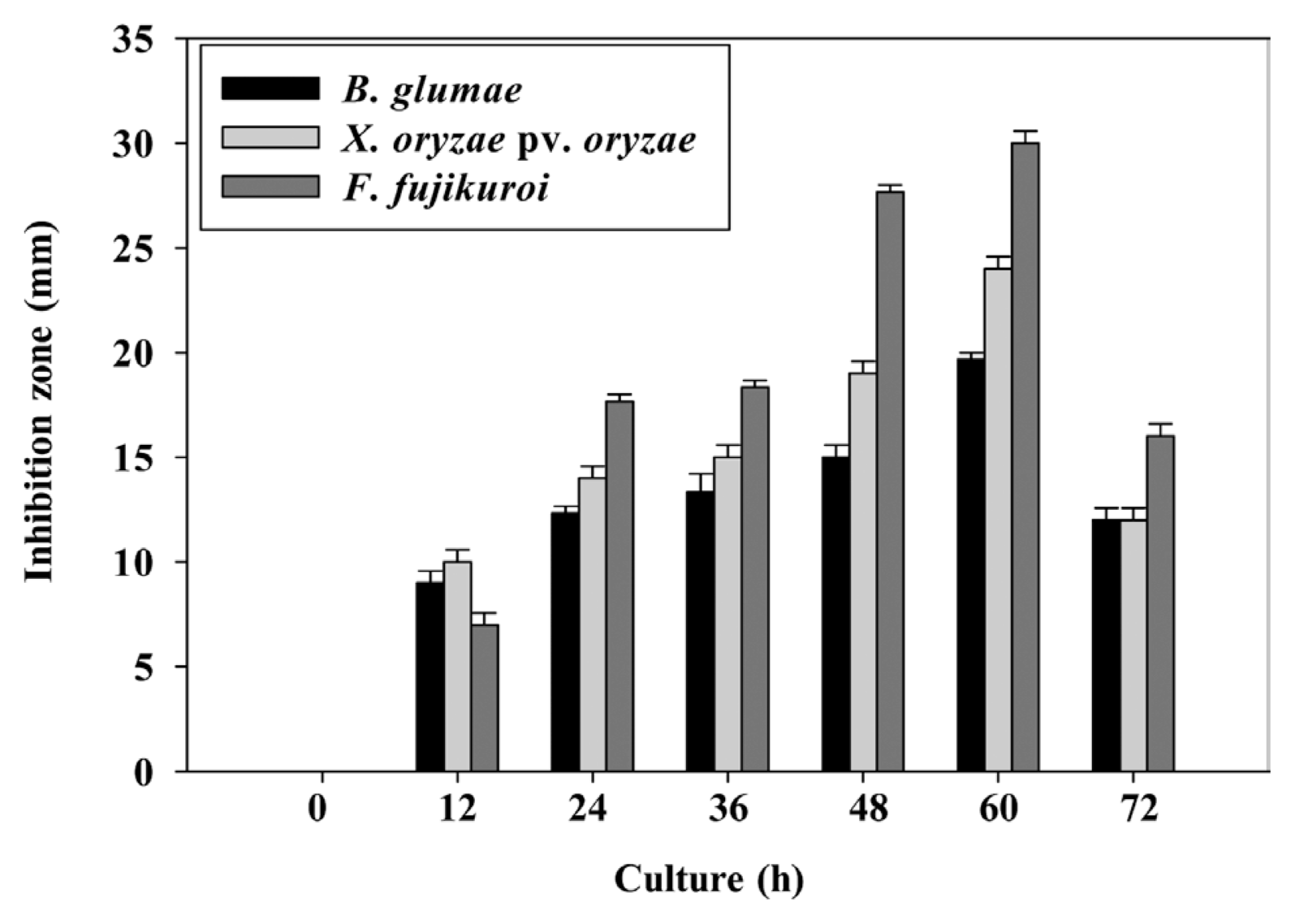
Among 250 bacterial strains isolated from the roots of rice plants, 15 endospore-forming bacteria showed inhibitory activity in the range of 2 to >20 mm to the mycelial growth of F. fujikuroi. These bacteria were found to be closely related to Paenibacillus polymyxa, Bacillus siamensis, Paenibacillus jamilae, Bacillus methylotrophicus, Bacillus thuringiensis, B. cereus, Bacillus simplex, and Bacillus daliensis, based on 16S rRNA gene sequences. These isolates were tentatively identified and their similarities were in the range of 99.27%-100% with the closely related genera. Of the isolates, two strains especially, YC7007 and YC7010T, with the highest similarity to B. siamensis showed strong antagonistic activity (>20 mm inhibition zone) against F. fujikuroi (Table 1). YC7007 also showed strong antagonistic activity against other rice pathogens, B. oryzae, M. grisea, F. fukikuroi, and other major plant pathogens, with inhibition zones from 10 to 29 mm on PDA (Table 2). The culture filtrate of strain YC7007 prepared from different cultivation times also showed good growth inhibition against F. fujikuroi and two major seed-borne bacterial pathogens of rice, B. glumae and X. oryzae pv. oryzae (Fig. 1). The culture filtrate of 60 h-old broth had the strongest activity against three pathogens on the respective culture media with the inhibition zone of 30 mm, 24 mm and 19.7 mm for F. fujikuroi, X. oryzae pv. oryzae and B. glumae, respectively.
Induction of systemic resistance against panicle blight and bacterial blight
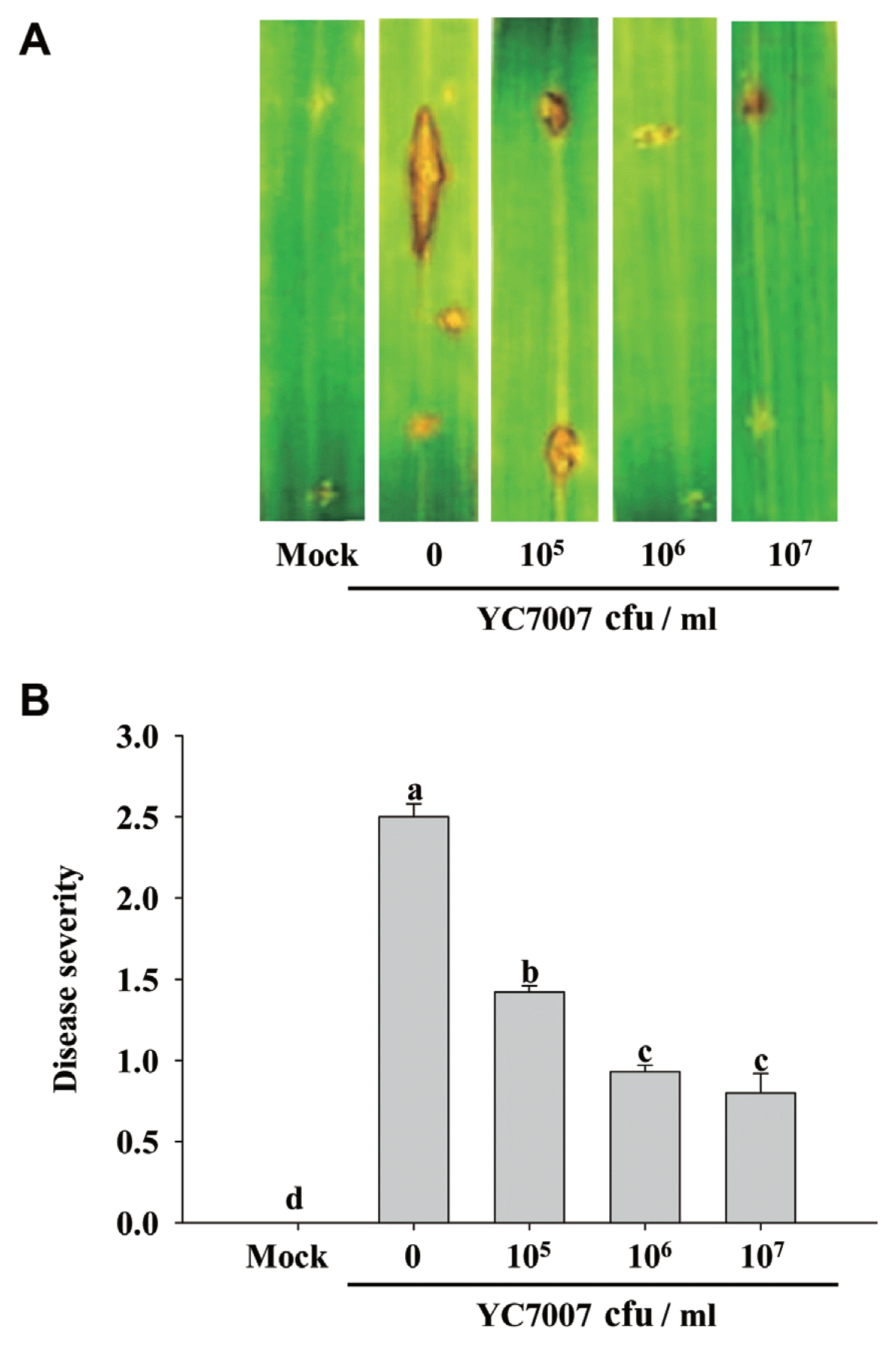
Strain YC7007 was investigated for the activity of inducing systemic resistance to the panicle blight and bacterial blight of rice. Drenching of three different concentrations of YC7007 to soils at 5.6├Ś105, 3.6├Ś106, and 2├Ś107 cfu/ml revealed significantly lower disease severity index of the leaf rot (p<0.05) by 1.4, 0.9, and 0.8, respectively, than the control, 2.5 (Fig. 2). However, there was no significant difference in the disease severity between bacterial concentrations of 106 and 107 cfu/ml. The concentration (2├Ś107, cfu/ml) of strain YC7007 drenched to the rhizosphere soils showed good control efficacy against panicle blight and bacterial blight, by 65.2% and 61.2%, respectively. Its culture filtrate sprayed to the leaves also showed significantly lower disease severity than the control, with disease reduction by 70.8% and 70.5% for panicle blight and bacterial blight, respectively (Table 3). The strain YC7007 showed significantly lower disease severity (p<0.01) against panicle blight and bacterial blight than the control.
Growth promotion by YC7007
Strain YC7007 had growth promoting activity with significantly (p<0.05) better growth than the control at all stages tested such as the seedling, tillering, and booting stages of rice. Treatment with the bacterial suspension (2├Ś107 cfu/ml) to the rhizosphere of the rice increased shoot length from 11.67 cm to 19.33 cm in the test tube at the seedling stage, 36.45 cm to 46.33 cm, and 55.33 cm to 61.0 cm in the tillering and booting stages, respectively. The length of primary roots increased from 2.67 cm to 7.67 cm at the seedling stage and the numbers of tillers were also increased, from 1.9 to 2.9 at the tillering stage and 5.3 to 7.0 at the booting stage (Table 4).
Identification of two antagonistic bacteria YC7007 and YC7010T
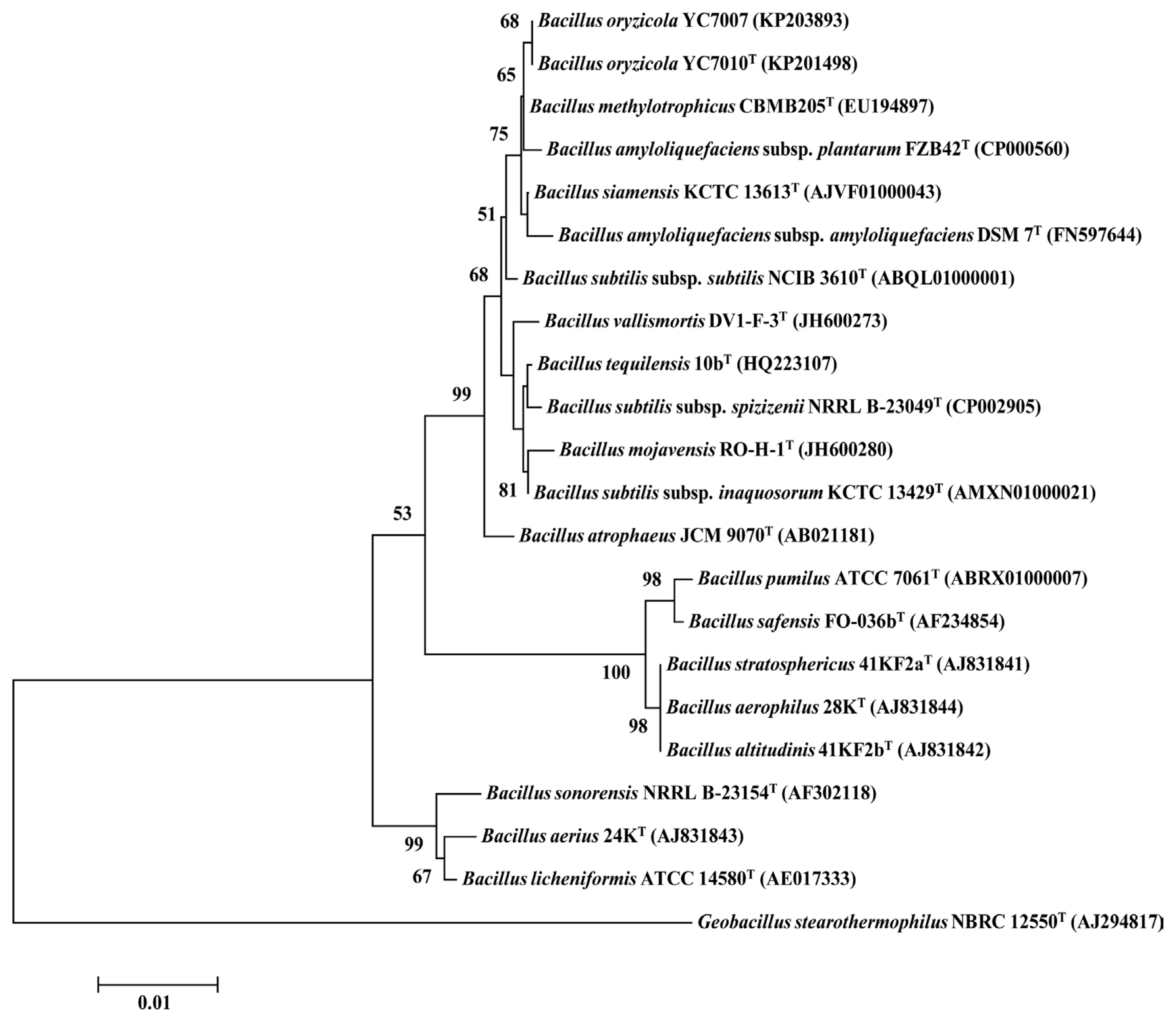
Two endophytic strains YC7007 and YC7010T with the strongest antagonistic activity out of 15 antagonistic bacteria were identified using a polyphasic approach. The 16S rRNA gene sequence of the two strains was a continuous stretch of 1513 base pairs, which aligned identically except only one nucleotide, with the database accession numbers KP203893 (YC7007) and KP201498 (YC7010T) in GenBank/EMBL/DDBJ. Based on the comparative 16S rRNA gene sequence analysis, the two strains are in the same clade with the same branch length of the phylogenetic tree, showing the highest similarity to B. siamensis KACC 15859T (99.67%), B. methylotrophicus KACC 13105T (99.65%), B. amyloliquefaciens subsp. plantarum KACC 17177T (99.60%), and B. tequilensis KACC 15944T (99.45%) (Table 1, Fig. 3). The DNA-DNA relatedness value between strain YC7010T and its most closely related strain B. siamensis KACC 15859T was 50.4┬▒3.5, but it was 91.5┬▒11.0% between YC7007 and YC7010T (Table 5). Both strains were Gram-positive, motile, rod-shaped, and grew well at 13-60┬░C (optimum at 28-30┬░C) and pH (4-12) (optimum at pH 7). Strains were able to grow at 0.1 TSA media with 1-13% NaCl (w/v), but NaCl was not required for growth. Strains were resistant to chloramphenicol by 30 ╬╝g/ml. Strains contained meso-diaminopimelic acid as the diagnostic diamino acid in the cell-wall peptidoglycan and MK-7 as a major respiratory quinone system. Other physiological and biochemical characteristics of two strains YC7007 and YC7010T are summarized in Table 6. The major fatty acids of the strains YC7007 and YC7010T were the anteiso-C15:0 (38.4 and 32.0%) and iso C15:0 (28.1 and 27.7%), respectively. The other cellular fatty acids profiles of YC7010T consisted of C16:0 (7.7%), iso C17:0 (6.4%), anteiso-C17:0 (5.3%), iso C16:0 (5.2%), C18:0 (5.1%), C16:1 Žē7c alcohol (3.4%), iso C14:0 (2.9%), iso-C17:1 Žē10c (1.7%), C16:1 Žē11c (1.4%), C14:0 (1.1%), and C20:1 Žē7c (0.2%) (Table 7). The G+C contents of the genomic DNA of two strains YC7007 and YC7010T were 50.5 mol% and 51.2 mol%, respectively. Strains exhibited a polar lipid profile consisting of phosphatidylethanolamine (PE), phosphatidylglycerol (PG), diphosphatidylglycerol (DPG), unknown glycolipid (GL) and unknown aminolipids (AL1-2) (Fig. S1). Based on 16S rRNA gene sequence analysis, DNA-DNA relatedness value, fatty acid composition and biochemical and physiological characteristics, strains YC7007 and YC7010T represent a novel species of the genus Bacillus, for which the name Bacillus oryzicola sp. nov. is proposed.
Description of Bacillus oryzicola YC7010T sp. nov
Bacillus oryzicola (o.ry.ziŌĆÖco.la L. fem. n. oryz, rice; L. v. colere, to inhabit; L. adj. oryzicola, living in rice). Cells are Gram-positive, rod-shaped (0.8-0.9├Ś2.0-3.0 ╬╝m). Colonies grown on R2A agar at 28┬░C for 2 days are white-cream, circular, and flat elevation with an entire margin. Cells are motile with a single polar flagellum. The cell wall contains meso-diaminopimelic acid as the diagnostic diamino acid. Cells usually occur singly or in pairs that are catalase-positive and oxidase-negative. Cells grow at 13-60┬░C and at pH 4-12.0. Hydrolysis of casein and gelatin is positive while negative for starch, Tween 20, Tween 80, tyrosine, and carboxymethyl cellulose. It uses D-glucose, D-fructose, D-mannose, D-mannitol, methyl-╬▒-D-glucopyranoside, N-acetyl-glucosamine, amygdalin, arbutin, esculin ferric citrate, salicin, D-celiobiose, D-maltose, D-lactose, D-saccharose, D-trehalose, D-raffinose, starch, glycogen, gentiobiose, trisodium citrate, and KohnŌĆÖs gelatin. With the API ZYM kit, it showed enzymatic activities of esterase (C4), esterase lipase (C8), naphthol-AS-Bl-phosphohydrolase, and N-acetyl-╬▓-glucosaminidase, but not lipase (C-14), leucine arylamidase, valine arylamidase, cystine arylamidase, trypsin, ╬▒-chymotrypsin, ╬▒-galactosidase, ╬▓-galactosidase, ╬▓-glucosidase, ╬▒-mannosidase, and ╬▒-fucosidase. It was resistant to 30 ╬╝g of chloramphenicol and streptomycin, but susceptible to 10 ╬╝g of ampicillin, penicillin, gentamycin, and 30 ╬╝g of kanamycin, vancomycin, and tetracycline. Growth was observed in R2A broth containing 13% (w/v) NaCl but not 14% (w/v). The major quinone is MK-7. The polar lipids are phosphatidylethanolamine (PE), phosphatidylglycerol (PG), diphosphatidylglycerol (DPG), unknown glycolipid and unknown aminolipids. The DNA G+C content of the type strain is 51.2 mol%. The type strain, YC7010T (= KACC18228T), was isolated from the root of rice in Jinju, Korea.
Discussion
Among many endophytic bacterial strains isolated from the rice roots, 6% of the Bacillus species were antagonistic and inhibited the mycelial growth of phytopathogens in this study. About 9% of endophytic bacteria including several Bacillus species and other Gram-negative bacteria isolated from tidal flat plants were antagonistic against oomycetous phytopathogens in a previous study (Bibi et al., 2012). Our strains, belonging to the genus Bacillus based on the 16S rRNA gene sequence, were examined for their antagonistic activity against the major rice fungal and bacterial pathogens in vitro and in vivo tests. Of these isolates, two strains YC7007 and YC7010T with the highest similarity to B. siamensis showed strong antagonistic activity in suppressing mycelial growth of the bakanae pathogen, F. fujikuroi, one of the most important seed-borne pathogens of rice. The culture filtrate of YC7007 also had strong inhibiting activity against bakanae, bacterial blight, and grain rot, suggesting that the strain may produce antifungal compounds. It has been reported that some Bacillus species produce small peptides and lipopeptides, such as fengycin, iturin, and surfactin, which have good suppression of phytopathogens (Bais et al., 2004; Crane et al., 2013; Dimkic et al., 2013). Drenching with a cell suspension of strain YC7007 suppressed the severity of panicle blight and bacterial blight of rice in pot tests, which indicated the induction of systemic resistance. Disease severity of panicle blight was reduced by treatment with the strain YC7007 at 105 (cfu/ml) and higher concentrations between 106 to 107 cfu/ml suppressed disease, over 62%. The culture filtrate of YC7007 also decreased panicle blight and bacterial blight significantly over 70% compared with the control. It was demonstrated that treatment with the cell suspension of B. subtilis (2.5├Ś108 cfu/ml) and B. cereus AR156 (5├Ś108 cfu/ml) controlled root and leaf speck diseases of Arabidopsis, respectively (Bais et al., 2004; Niu et al., 2011). The bacterial concentrations of these two isolates B. subtilis and B. cereus AR156 (108 cfu/ml) was over 100 times higher than that of strain YC7007 (106 cfu/ml), which may not be feasible for practical use at commercial farms. For developing antagonistic bacteria as biological agents, the optimal concentration of bacteria should be equal or lower than 107 cfu/ml, considering the dilution factor in the final use by farmers (Chen et al., 2009). As the bacterial suspension drenched to the rhizosphere of rice and culture filtrate sprayed on the leaves, had an antagonistic activity against the pathogens, strain YC7007 probably has indirect and/or direct interactions between them. Diseases of panicle blight and bacterial blight were controlled by the treatment - both drenching and spraying - suggesting that the strain YC7007 may react through induced systemic resistance (ISR) or systemic acquired resistance (SAR) for the suppression of the diseases (Ahn et al., 2007; Niu et al., 2011). The reaction mechanism needs to be determined in terms of the hormonal signaling via the salicylic, jasmonic, or ethylene pathways for SAR or ISR.
Several species of Bacillus have been commercialized as biological agents due to their long persistence in adverse environments over a long period. Some of these were functional in many plants, by inducing systemic resistance (Hu et al., 2011; Kloepper et al., 2004). B. vallismortis EXTN-1 and B. cereus have been used to control rice blast, sheath blight, and bakanae (Kazempour and Elahinia, 2007; Park et al., 2006). To our knowledge, this is the first report that Bacillus strain YC7007 has long-lasting activity in resistance induction in rice against major bacterial diseases of panicle blight and bacterial blight. Furthermore, strain YC7007 showed good growth promotion of rice by a one-time application of a bacterial suspension at the seedling stage, just after germination. YC7007 increased shoot and root length by 1.1-2.9 fold at the seedling, tillering, and booting stages, and increased tiller numbers by over 1.3-fold at the tillering and booting stages compared with the control. This demonstrates that resistance-inducing and growth-promoting activity by the probiotic bacterium YC7007 could be long lasting, from the seedling to the booting stage (Picard et al., 2008). It has been reported that the induction of host resistance negatively affects host plants, in suppressing growth associated with the plant hormones, gibberellic acid (GA) and auxin. The immune system is mediated by brassinosteroids and salicylic acid, repressing GA and auxin responses, at least in part via negative cross talk with the hormone, respectively, in rice and Arabidopsis (De Vleesschauwer et al., 2012; Wang et al., 2007). A chemical derivative of salicylic acid (SA), benzothiadiazole (BTH), induced systemic resistance against biotrophic pathogens in rice, cucumber, Arabidopsis, and pepper plants via salicylic acid signaling networks, but it retarded the plant growth by inhibiting the auxin response. In contrast, some rhizobacteria, including Bacillus species, enhanced resistance induction with no negative effect on plant growth (Ahn et al., 2005; Ryu et al., 2004; Yang et al., 2009). In this respect, strain YC7007 may be a good candidate for developing a biocontrol agent with multiple functions of resistance induction and growth promotion in rice without negative effect. Additionally, strain YC7007 has anti-fungal and anti-bacterial activity, probably via production of antibiotics.
Our two endophytic strains, designated YC7007 and YC7010T, were further characterized based on a polyphasic study including 16S rRNA gene sequences, DNA-DNA hybridization, fatty acid analyses, and other physical and biochemical tests. The two strains were identified as novel species of the genus Bacillus, in the same clade with the same branch length in the phylogenetic tree. The overall percentage sequence divergences of the two strains were the same, but differed from the other closely related species in the clade of the phylogenetic tree. Strains YC7007 and YC7010T showed 100% similarity and very high DNA-DNA relatedness values of 91.5% between them, indicating that the two strains are the same Bacillus species. The DNA-DNA hybridization values of other related strains B. siamensis, B. methylotrophicus, B. subtilis subsp. inaquosorum, B. amyloliquefaciens subsp. plantarum, and B. tequilensis in comparison to the type strain YC7010T were below 70%, which indicated that YC7010T was a novel species (Goris et al., 2007; Stackebrandt and Goebel, 1994). Additionally, the two strains showed almost the same reactions in physical and biochemical tests using API kits, but showed different responses to other reference strains, indicating that the two strains are distinct from other related Bacillus strains. In tolerance to NaCl, the two strains can survive up to 13% NaCl, but B. methylotrophicus did not grow at 10% NaCl (Madhaiyan et al., 2010). Major cellular fatty acids of YC7007 and YC7010T were anteiso-C15:0 and iso C15:0, which are same as other related species of the genus Bacillus. Other minor fatty acids of the two strains differed from those of closely related Bacillus strains. Each bacterium has a specific fatty acid profile and fatty acid analysis is used as an effective tool in bacterial taxonomy (K├żmpfer, 1994). The major isoprenoid quinones of strains YC7007 and YC7010T were menaquinone-7 (MK-7) as those of the type strains of the genus Bacillus species (Kang et al., 2013). The DNA G+C contents of both strains were 50.5-51.2 mol%, which are similar to but slightly higher than those of other Bacillus species (Madhaiyan et al., 2010; Sumpavapol et al., 2010). Major polar lipids of both strains were PE, PG, and DPG, which are in accordance with those of B. siamensis and B. songklensis (Kang et al., 2013; Sumpavapol et al., 2010).
Based on all the data from the polyphasic studies, the two strains YC7007 and YC7010T, belonged to the genus Bacillus; we therefore, proposed strain YC7010T and YC7007 as a novel species and suggested the name Bacillus oryzicola YC7010T sp. nov. with the type strain. In conclusion, the endophytic strain B. oryzicola YC7007 has multifunctional activities in rice: direct inhibition of fungal and bacterial pathogens, systemic resistance induction, and plant growth promotion, which could possibly be used practically as a microbial inoculant on commercial farms.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






