

Pepper (Capsicum annuum L.) is an economically very important crop. In the last decade, production of pepper increased by 40% (Kim et al., 2014). South Korea produced about 117,800 tons of pepper from approximately 45,400 hectares in 2013 (KOSTAT, 2013). Fungal species in the genus Colletotrichum cause anthracnose on many vegetables and fruits, including peppers. Anthracnose has been responsible for an estimated 10% annual reduction in pepper productivity, equivalent to about US $100 million, in South Korea (Kim et al., 2008). In the 1990s, anthracnose symptoms on pepper was reported to be caused by C. gloeosporioides, C. dematium, and C. coccodes, with C. gloeosporioides representing up to 90% of the Colletotrichum spp. field population (Park and Kim, 1992). Since that time, a more aggressive form of anthracnose disease, occurring at all stages of pepper development, has reportedly been caused by C. acutatum. Colletotrichum acutatum can also infect other crops, including tomatoes (Solanum lycopersicum L.), strawberries (Fragaria ananarsa), and apples (Malus domestica), whereas C. gloeosporioides has frequently been associated with anthracnose on other fruit and vegetable crops including apple, tangerine (Citrus tangerine), persimmon (Diospyros spp.), grape (Vitis spp.), strawberry, watermelon (Citrullus lanatus), and ginseng (Panax spp.) in South Korea (Kim and Nam, 1999).
Anthracnose is usually controlled with application of chemical fungicides such as benomyl, maneb, chlorothalonil, and mancozeb (Mahoney and Tattar, 1980). Increased use of chemical fungicides has led to the development of fungicide-resistant fungal strains (Howarth and Francis 1991; Peres et al., 2004). In particular, C. acutatum is resistant to benomyl and other benzimidazole fungicides (Peres et al., 2004). Molecular analysis revealed that amino acid substitutions in β-tubulin conferred this resistance to the benzimidazole fungicides (Jung and Oakley, 1990). The use of pesticides continues to increase public concerns over risks associated with hazardous residues on agricultural products (Jang et al., 2010), long-term and unknown effects on health (Miles and Frewer, 2001), and adverse effects on biotic diversity in ecosystems (Kegley et al., 1999). Disease control by pesticide application is unaffordable in developing countries (Wesseling et al., 1997). For these reasons, biological control using antagonistic microorganisms has emerged as an environmentally friendly alternative for plant pathogen control (Pal and Gardener, 2006).
Antagonistic microorganisms produce a wide variety of antimicrobial metabolites (Keel et al., 1992; Ongena and Jacques, 2008; Raaijmakers et al., 2002). Plant growth promoting rhizobacteria (PGPR), a group of plant-beneficial bacteria in the rhizosphere, are antagonistic to plant pathogens by producing secondary metabolites, including siderophores, lytic enzymes, antibiotics, and cyanide (Chet et al., 1990; Than et al., 2004). Although PGPR properties responsible for plant growth promotion have not been fully elucidated, PGPR is known to produce plant growth regulators such as indole-3-acetic acid (IAA) and promote asymbiotic N fixation and solubilization of soil P compounds (Idris et al., 2007; Malik et al., 1997; Rodriguez and Fraga, 1999). Antagonistic microorganisms include Bacillus spp., Pseudomonas spp., Paenibacillus spp., and Streptomyces spp., which are mainly isolated to control the pepper anthracnose from the rhizosphere and plant microflora (Garbeva et al., 2004; Jeon et al., 2010; Lamsal et al., 2012; Lee et al., 2011; Lim, 2005; Weller, 2007; Yoo et al., 2012). However, there is wide range of environments where more beneficial microorganisms have not been discovered.
Tidal flats on coastlines and on the shores of lagoons and estuaries in intertidal areas are highly productive, as they support microbial growth with an abundance of organic matter (Kim et al., 2004). We isolate antagonistic bacterial strains from tidal flats and determine their antagonistic properties by investigating the production of compounds including siderophores, proteases, and chitinases, and their ability to solubilize insoluble phosphatase. In addition, antagonistic activities of selected bacteria were evaluated in the presence of fungal pathogens C. acutatum and C. gloeosporioides by measuring suppression of growth, germination, and disease severity.
Materials and Methods
Isolation of bacterial strains
Soil samples were collected from mudflats on the west coast of Youngjongdo (37° N. lat., 126° E. long) in South Korea, placed in sterilized polythene bags, brought to the laboratory, and stored at 4°C. To isolate bacterial strains from the mudflats, 1 g of mudflat sample was mixed with 9 ml sterile water. The mudflat suspensions were serially diluted with sterile water, after which a 1 ml 10−5 dilution was plated on LB agar (trytone 1%, yeast extract 0.5%, NaCl 0.5%, agar 1.5%) and marine agar (Difco, Detroit, MI, USA) plates. The plates were incubated at 28°C for 3 days, after which colonies were sub-cultured onto a new plate of the same media to isolate single colonies. The isolated bacterial strains were stored in LB containing 20% glycerol at −70°C.
Identification of bacterial isolates
DNA was extracted from the bacterial cells by harvesting 3 ml cells following overnight culture. Chromosomal DNA was extracted using a Wizard genomic DNA purification kit (Promega, USA). The 16S rRNA was amplified with the universal primers 27F (AGAGTTTGATCMTGGCTCAG), 1492R (TACGGYTACCTTGTTACGACTT), in Thermal Cycler Thermal Controller 2720 (Applied Biosystems, USA). PCR products were purified using a fragment DNA purification kit (Intron, Korea), and were subsequently sequenced using primers 518F (CCAGCAGCCGCGGTAATACG) and 800R (TACCAGGGTATCTAATCC; Macrogen Inc., Seoul, Korea). The sequences were blasted against the BLASTN program of NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi). DNA sequences from the NCBI nucleotide database were aligned with the ClustalW program in MEGA 6.0, and the phylogenetic relationship was constructed using the nearest-neighbor data analysis method with 1,000 bootstrap replicates.
Assays for inhibition of hyphal growth and conidial germination
Antifungal activity was imposed on a PDA plate with holes (4 mm in diameter) punched using a cork borer. Bacterial strains were inoculated 1 day before inoculation of the fungal pathogens, and 20 μl bacteria culture were dropped into the holes. All of the cultures were incubated at 25°C in the dark. The growth of fungal pathogens was evaluated 10 days after incubation. Inhibition of fungal growth was measured as the distance of the clear zone between the bacterial colony and each fungus according to the following formula:
where TD and CD were the distance between the center and fungal hyphae edges in the treatment and control plates, respectively. Each experiment was replicated three times, and three separate experiments were performed.
For conidial germination assay, Collectorichum species were incubated on PDA plates at 25°C, and conidia production was induced. After 10 days, conidia were harvested with distilled water. Conidial suspension concentration was adjusted to 1×104 conidia/ml using a hemocytometer. Each bacterial culture (OD 1.0) diluted serially (10, 100, and 1000 times) was co-mixed with the conidial suspension, placed on a hydrophobic slide (Knittel Glaser, Germany), and incubated at 25°C in a humidity box for 20 h. Inhibition rate of conidial germination was calculated, according to the following formula (Kwak et al., 2012):
where T represented germination rate of the treatment, and C represented germination rate of the control. Each experiment was replicated three times, and three separate experiments were conducted.
Analyses of antimicrobial and antagonistic substances
Chitin degrading ability was tested in colloidal chitin agar media containing 0.7 g/l K2HPO4, 0.3 g/l KH2PO4, 0.5 g/l MgSO4·5H2O, 0.01 g/l FeSO4·7H2O, 0.001 g/l ZnSO4, 0.001 g/l MnCl2, 15 g/l agar, and 0.5 g colloidal chitin dissolved in 1 l distilled water (Hsu and Lockwood, 1975). Colloidal chitin was prepared as follows. Chitin powder 80 g was dissolved in 1 L concentrated hydrochloric acid and continuously stirred on ice for 1 h. After stirring, the hydrolyzed chitin was washed several times with distilled water to remove the acid, using filter paper, after which pH was adjusted to a range of 6-7. This colloidal chitin was stored as a paste at 4°C. After incubation for 3 days at 25°C, development of a clear zone indicated chitinase activity.
Proteolytic activity was tested by LB agar plates containing with 3% skim milk powder (Sokol et al., 1979). After 3 days incubation at 28°C, clear zone surrounding the bacterial suggested protease production.
Screening for cellulase producers was conducted on CMC agar (1.0 g/l KH2PO4, 0.5 g/l MgSO4·7H2O, 0.5 g/l NaCl, 0.01 g/l FeSO4·7H2O, 0.01 g/l MnSO4·H2O, 0.3 g NH4NO3, 10 g/l carboxymethyl cellulose, 12 g/l agar) media (Teather and Wood, 1982). After incubation at 25°C for 3 days, the agar was flooded with 0.1% Congo red for 15 min to 20 min, and then with 1 M NaCl for 15 min to 20 min. After staining, agar plates were observed for zones around the colonies.
Phosphate solubilization screening was performed on Pikovskaya’s media (Nautiyal, 1999) containing 10 g/l glucose; 5 g/l Ca3(PO4)2, 0.5 g/l (NH4)2SO4, 0.2 g/l NaCl, 0.1 g/l MgSO4·7H2O, 0.1 g/l KCl, 0.2 g/l yeast extract, 0.001 g/l MnSO4·H2O, and 0.001 g/l FeSO4·7H2O. The pH of the media was adjusted to 6.8 before autoclaving. After incubation for 15 days at 25°C, the development of a clear zone indicated phosphate solubilization.
Siderophore production was measured using the modified Chrome azurol S (CAS) agar assay (Schwyn and Neilands, 1987). CAS agar plate was prepared as follows. A total of 60 mg CAS was dissolved in 50 ml deionized water and mixed with 10 ml iron (III) solution (2.7 mg FeCl3·6H2O, 10 mM HCl). This solution was slowly mixed with 73 mg HDTMA dissolved in 40 ml water. The resulting blue dye solution was autoclaved and mixed with a solution of 750 ml, 15 g/l agar, 32.24 g/l PIPES, and 100 ml MM9 salt stock solution (30 g/l KH2PO4, 50 g/l NaCl, 100 g/l NH4Cl in 1 l ddH2O). This solution was autoclaved and cooled to 50°C, to which we slowly added 100 ml blue dye, 30 ml 10% Casamino acid solution, and 10 ml 20% glucose solution along the glass wall, with sufficient agitation to mix thoroughly. The modified CAS agar plate was punched with 4 mm diameter holes using a cork borer. Each hole was filled with a 20 μl of bacterial suspension. LB broth was used as the negative control. Siderophore production was measured after 5 days of incubation at 25°C.
Disease suppression assay
For the pathogenicity assay, pepper fruits were surface sterilized for 1 min in 1% NaOCl, briefly rinsed with distilled water, and air-dried at room temperature. Each bacterial culture suspension (1.0 O.D) was sprayed. After 24 h, a C. acutatum spore suspension (3.5×104 conidia/ml) was inoculated on pepper fruits, and incubated at room temperature in a humidity box. After 2 weeks, the diseased areas of pepper fruits were measured visually and by image assessment with computer programs (ImageJ 1.48 V).
Biochemical and physiological characteristics
Analytical profile indices (APIs) from kit API 50CHB systems (BioMérieux, Marcy l’Etoile, France) were used to characterize the physiological and biochemical properties of the bacterial isolates. API panel test system substrate utilization tests were performed using the API 50CHB systems panel. API test strips were handled according to the manufacturer’s instructions (bioMerieux, Marcy l’Etoile, France). Stock cultures were streaked onto LB agar to obtain single colonies for each bacterial isolate. Bacterial colonies of each isolate were diluted in 0.85% NaCl solution. The amount of bacteria was adjusted to 1 McFarland, and 200 ml of this solution was transferred into each well of the panels. To prevent contamination from the air, the wells were filled with mineral oil, and incubated at 25°C. Readings were recorded after 48 h.
Results
Antifungal activity and identification of bacterial strains
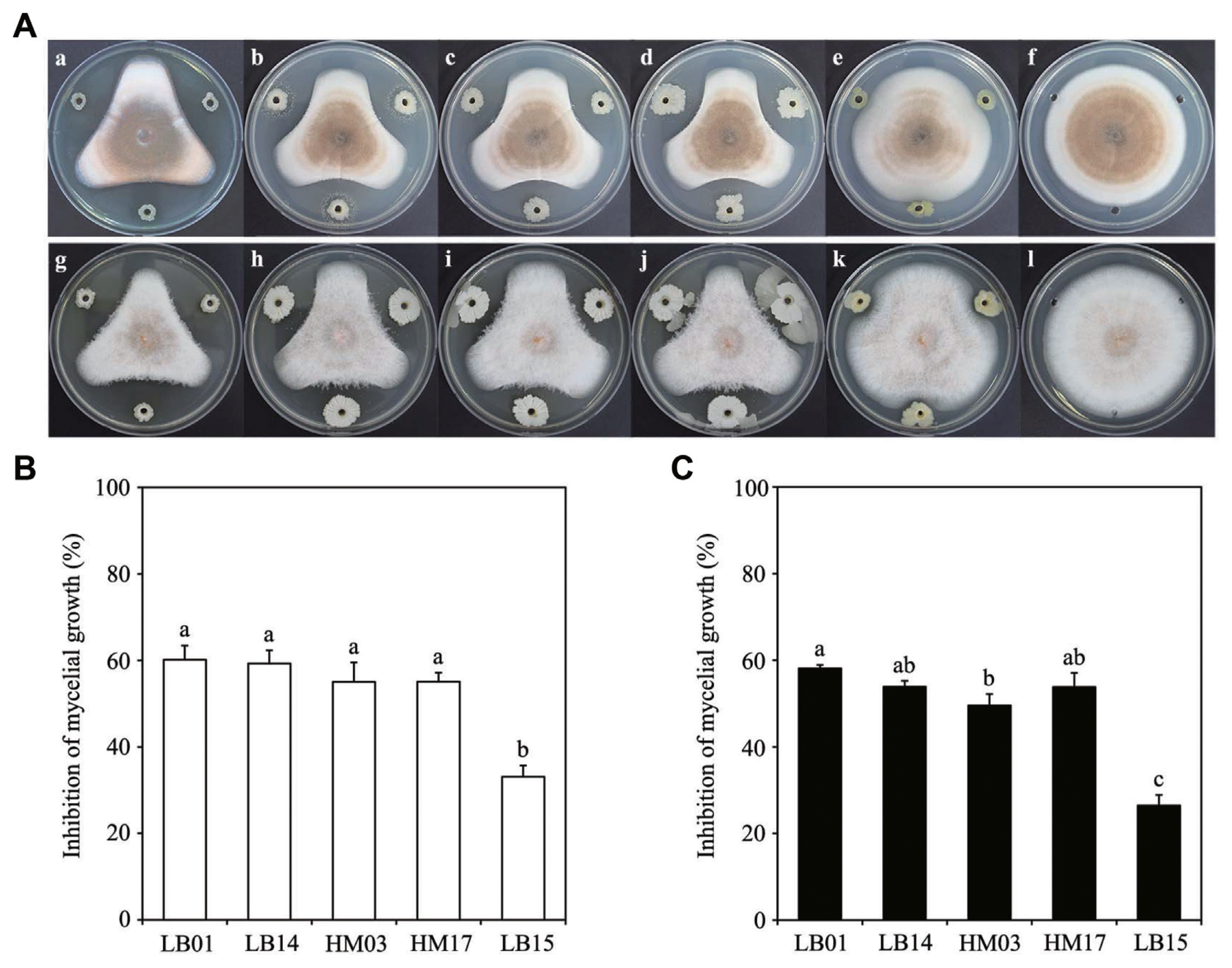
Soil samples from mudflats of the western sea of South Korea were used to isolate bacteria with antifungal properties. A total of 420 bacteria species were isolated and categorized into 13 groups, based on colony morphology and pigmentation. A representative member of each group was screened for antifungal activity against C. acutatum and C. gloeosporioides using a dual culture method.
As shown in Fig. 1A, five bacteria strains inhibited growth of C. acutatum and C. gloeosporioides. The strain LB01 exerted the highest inhibition of mycelial growth on C. acutatum (60.15%) and C. gloeosporioides (58.12%) compared to the control (Fig. 1B, C). The strain LB14 showed 59.29% and 53.87% inhibition of mycelial growth of C. acutatum and C. gloeosporioides, respectively. HM03 and HM17 also inhibited mycelial growth of C. acutatum and C. gloeosporioides by more than 50%. The strain LB15 was the least effective among the five strains in inhibiting mycelial growth of the two fungal species.
To identify the five strains with antifungal activity, their 16S rRNA sequences were analyzed. Sequence data revealed that the five bacterial isolates belonged to the Bacillus or Staphylococcus genus (Table 1). The three strains, LB14, HM03, and HM17, were distantly related and grouped with B. atrophaeus. The two strains, LB01 and LB15, were very closely grouped with B. amyloliquefaciens and B. pumilus, respectively (Fig. 2).
Production of antimicrobial and antagonistic substances
To examine antifungal activity of the isolated Bacillus species, they were tested for production of antimicrobial substances, including chitinase, protease, and cellulase, on LB plates containing colloidal chitin, skim milk, or carboxymethyl cellulose (Fig. 3A). Three strains, LB14, HM03, and HM17, were able to decompose chitin, but LB01 and LB15 were unable to degrade chitin. Strong proteolytic activity was found in LB01, LB14, HM03, HM17, and LB01. LB15 formed a very faint zone, suggesting less proteolytic activity. Strong cellulase activity was observed for LB01. LB14, HM03, and HM17 had no cellulase activity.
As a next step, we determined the contribution of siderophore production and solubilization of insoluble phosphate to antagonistic properties of the isolated Bacillus species (Fig. 3B). All of the tested strains formed halo zones on CAS agar media, indicating strong siderophore production. Among the five strains, LB14, HM03, and LB15 had relatively higher siderophore production. The highest solubilization capacity of insoluble phosphate was found for LB14, HM03, and HM17.
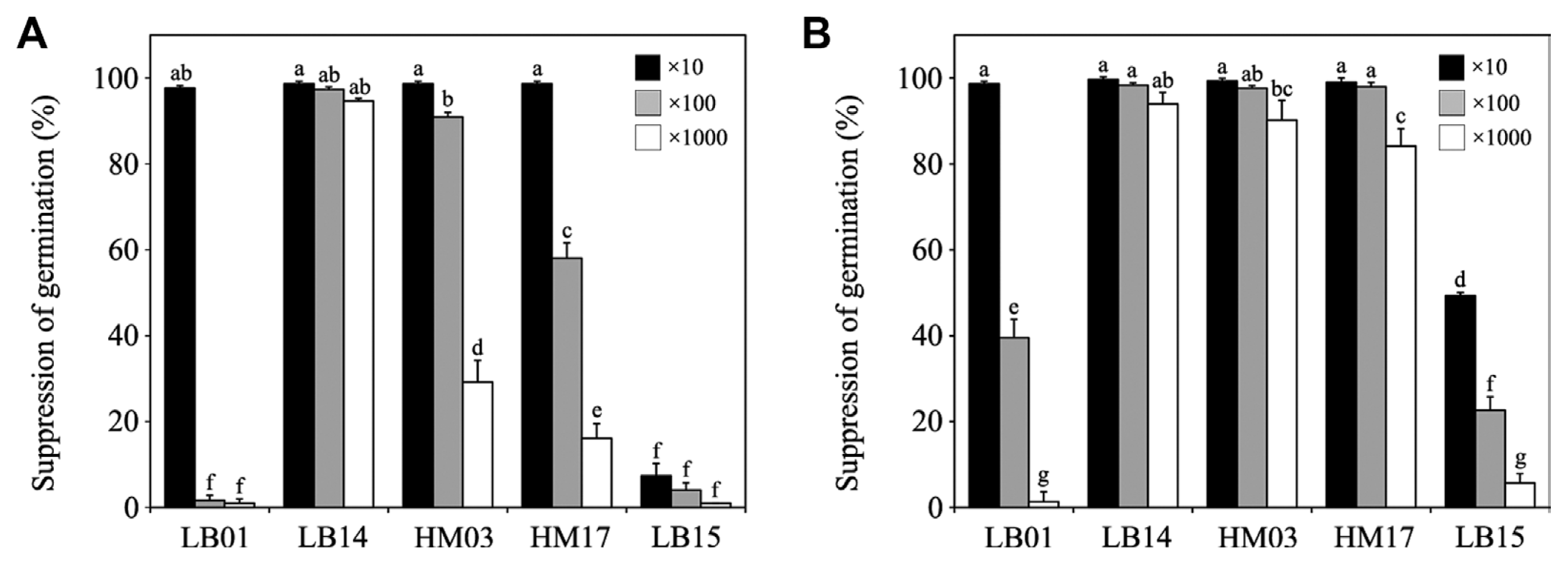
Suppression of conidial germination of C. acutatum and C. gloeosporioides by Bacillus strains
Because conidial germination is an early stage of disease development, the ability of Bacillus strains to suppress conidial germination of C. acutatum and C. gloeosporioides was investigated (Fig. 4). Ten-fold diluted cultures of the other four strains, LB01, LB14, HM03, and HM17, highly suppressed conidial germination of C. acutatum (Fig. 4A) and C. gloeosporioides (Fig. 4B). Cultures of LB15 were not effective for suppressing conidial germination of C. acutatum, although the culture at a 10-fold dilution exhibited increased effectiveness on suppression of conidial germination of C. gloeosporioides. All of the serial dilution cultures of LB14 strongly inhibited conidial germination of both C. acutatum and C. gloeosporioides. HM03 cultures diluted to 10-fold and 100-fold effectively inhibited conidial germination of the two fungal isolates. Effectiveness of the HM03 culture diluted 1000-fold dropped to 29% in inhibiting conidial germination of C. acutatum, but it was stable in inhibiting germination of C. gloeosporioides. Clear differences were found in the ability of serial dilutions of HM17 cultures to inhibit conidial germination of C. acutatum compared to C. gloeosporioides.
We also tested the ability of Bacillus strains to suppress C. acutatum-mediated disease development on peppers. As shown in Fig. S1 and Table 2, diseased areas on peppers were significantly reduced with treatment of Bacillus strains. Diseased areas ranged from 2.9% to 45.9% on Bacillus-treated peppers, whereas the diseased area occupied 88.6% of Bacillus-untreated peppers (control). The highest suppression of pepper anthracnose disease was found on LB14-treated peppers with 96.7 % control efficiency.
Biochemical and physiological characterization of the LB14 strain
Based on antagonistic effects and the ability to suppress anthracnose, LB14 was selected for biochemical and physiological characterization (Table 3). This analysis revealed that LB14, identified as B. atrophaeus, was able to fully metabolize glycerol, l-arabinose, ribose, d-glucose, d-fructose, d-mannose, d-manitol, l-sorbitol, arbutin, esculin, salicin, cellobiose, sucrose, and trehalose. Unlike B. subtilis, LB14 was unable to metabolize d-xylose, inulin, and gluconate, suggesting that LB14 differs from B. subtilis in utilization of carbohydrates.
Discussion
In recent years, there has been increasing demand for safe and eco-friendly agricultural products (Huh and Kim, 2010). Accordingly, biological control practices are in much greater demand as alternatives to synthetic pesticides. Biological control practices are particularly important in organic crop production disease control (Cook, 1993; Han et al., 2013; Kim and Yun, 2011; Yang et al., 2002).
Reports from several studies provide strong evidence that strains of Bacillus species, including B. subtilis (Kim et al., 2012; Lee et al., 2006; Lee et al., 2011), B. amyloliquefaciens (Kong et al., 2010a; Lee et al., 2013; Wu et al., 2007), B. licheniformis (Govender et al., 2005; Kim et al., 2007; Kong et al., 2010b), B. pumilus (Akhtar and Siddiqui, 2008; Sari et al., 2007), B. mycoides, and B. sphaericus, significantly reduce disease severity on a variety of hosts in greenhouse or field conditions (Kloepper et al., 2004). B. subtilis and B. amyloliquefaciens have been used in commercial biological control products due to their excellent antagonistic effects and high stability in harsh environmental conditions (Kwak et al., 2012).
In the present study, we isolated antagonistic microorganisms from tidal flat sediments, including three strains of B. atrophaeus (LB14, HM03, and HM17, one strain of B. amyloliquefaciens (LB01), and one strain of B. pumilus (LB15). A previous study used tidal flat sediments and jeotgal, a salted-fermented fish product, to find antagonistic microorganisms in Korea. The antagonistic bacteria included Paenibacillus macerans, B. atrophaeus, and B. pumilus from tidal flat sediments, and B. atrophaeus, Paenibacillus sp. Virgibacillus pantothenticus, B. subtilis, and other Bacillus sp. from jeotgal (Kim et al., 2010).
Consistent with previous reports that B. atrophaeus suppresses fungal plant pathogens (Kim et al., 2010), our study demonstrated that three B. atrophaeus strains, LB14, HM03, and HM17, were effective for not only inhibiting mycelial growth, but also inhibiting conidial germination of C. acutatum and C. gloeosporioides. Furthermore, we showed that chinase, protease, siderophore, and phosphate solubilization, associated with LB14, HM03, and HM17, may contribute to suppression of C. acutatum and C. gloeosporioides.
Fungal cell walls play important physiological roles, in addition to providing structural barriers, in regulating the passage of molecules into cells and their development and survival in response to ambient conditions. The fungal cell wall is a matrix composed of polysaccharides, small amounts of proteins, and other components. Many fungal pathogens in ascomycota and basidiomycota have a mixture of chitin, polymers of glucan, and mannoproteins in their cell walls (Vega and Kalkum, 2012). Many antagonistic bacteria secrete lytic enzymes capable of dissolving fungal cell walls, resulting in exhibition of predatory activity of fungal pathogens (Xu et al., 2014). Strong activity of chitinase and protease in the B. atrophaeus strains LB14, HM04, and HM17 are therefore consistent with the observed growth inhibition of C. acutatum and C. gloeosporioides (Fig. 3A).
Proteases, in particular, play a key role in the cell lysis process. Proteases bind to the outer mannoprotein layer of the cell wall, open the protein structure, and expose inner glucan layers and chitin microfibrils. Among the three B. atrophaeus strains, LB01 (B. amyloliquefaciens) exhibited strong protease activity. However, activity of chitinase and protease was almost absent in LB15 (B. pumulis). This difference may explain the reduced effectiveness of LB15 compared to the other B. atrophaeus and B. amyloliquefaciens strains in inhibiting mycelial growth and conidial germination of C. acutatum and C. gloeosporioides.
The three B. atrophaeus strains, together with B. amyloliquefaciens and B. pumilus strains isolated from tidal flat sediments, demonstrated a high capacity to solubilize an insoluble phosphate compound. This is an important trait of antagonistic bacteria in supplying phosphate, a major nutrient required for plant growth. Uptake of phosphate by plants is limited due to low solubility of phosphate compounds in soil. Many bacteria are known to increase solubilization of insoluble phosphate forms by releasing organic acids and phosphatase enzymes (Halder et al., 1990; Rossolini et al., 1998; Sahu and Jana, 2000). Therefore, phosphate solubilizing bacteria, especially those with antifungal activities, may promote crop productivity, by not only providing plant-absorbable forms of phosphate, but also by effectively protecting plants from fungal soil-borne diseases (Dey et al., 2004; Sundara et al., 2002).
In addition, production of siderophore by the three B. atrophaeus strains may be a key factor in promoting plant growth and protecting plants from plant pathogens. Iron, an essential cofactor for cellular processes, is abundant in nature, but iron bioavailability is very limited in soils due to low solubility under aerobic conditions and in the presence of a neutral pH. A number of bacteria, including plant pathogens, produce iron-chelating siderophores. Siderophores produced by antagonistic microorganisms may inhibit the growth of plant pathogens, but may enhance plant growth by increasing iron in the root zone. Studies have demonstrated that beneficial bacteria-producing siderophores stimulate plant growth and inhibit germination of a soil-borne fungal pathogen (Alexander and Zuberer, 1991; Elad and Baker, 1985; Kloepper et al., 1980). Similarly, the variable efficiency in inhibiting conidial germination of C. acutatum and C. gloeosporioides may be correlated with siderophore production by the B. atrophaeus and B. pumilus strains, although the effect of strong antifungal substances produced by B. atrophaeus strains should also be considered (Fig. 3). The B. atrophaeus strain LB14 exhibited a strong antagonistic activity and was most effective for suppressing C. acutaum-mediated disease development on pepper plants compared to the other strains (Fig. S1 and Table 2), suggesting that it may be a promising agent for biological control and plant growth promotion. Further evaluation of LB14 should be performed under different environmental conditions, cultural practices, and fungicide applications, to obtain more knowledge of efficacy in the field. Knowledge of the mechanisms and performance of antagonistic microorganisms will be helpful in developing reliable biological systems for disease control.


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






