



Root-knot nematode (Meloidogyne spp.; RKN), the sedentary endoparasitic nematode, is one of the most economically significant plant-parasitic nematodes, more than 100 species were recognized in this genus (Janati et al., 2018). Symptoms associated with RKN infection can be easily diagnosed by farmers by the distinctive galls. Plants infected by RKN show symptoms such as; yellow leaves, stunted growth, nutrient deficiencies, and secondary infections by other pathogens (Hunt and Handoo, 2009; Mantelin et al., 2017). Approximately 29 species of nematodes have been described in association with losses in irrigated rice. Among of them, Paratrichodorus minor, Aphelenchoides besseyi, Ditylenchus angustus, Heterodera elachista, Hirschmanniella oryzae, Meloidogyne incognita, M. javanica, M. arenaria, and M. oryzae have significant detrimental effects (Mantelin et al., 2017; Negretti et al., 2017). Meloidogyne graminicola, one of the major species in RKN, was first isolated from the roots of barnyard grass (Echinochloa colonum L.) in Baton Rouge, LA, USA, and was reported by Golden and Birchfield (1965). Subsequently, this nematode was identified in many countries. The nematode has a wide range of alternative hosts; cereals, dicotyledonous, grass plants, and rice, the most important host (Bridge et al., 2005; Rich et al., 2009). M. graminicola is the most prevalent plant-parasitic nematode on rice and is considered a major threat to rice. This nematode cause 16-20% loss in lowland rainfed rice in Bangladesh, and cause 16-32% yield losses under irrigated and 11-73% under flooded conditions in India, respectively (Tian et al., 2018).
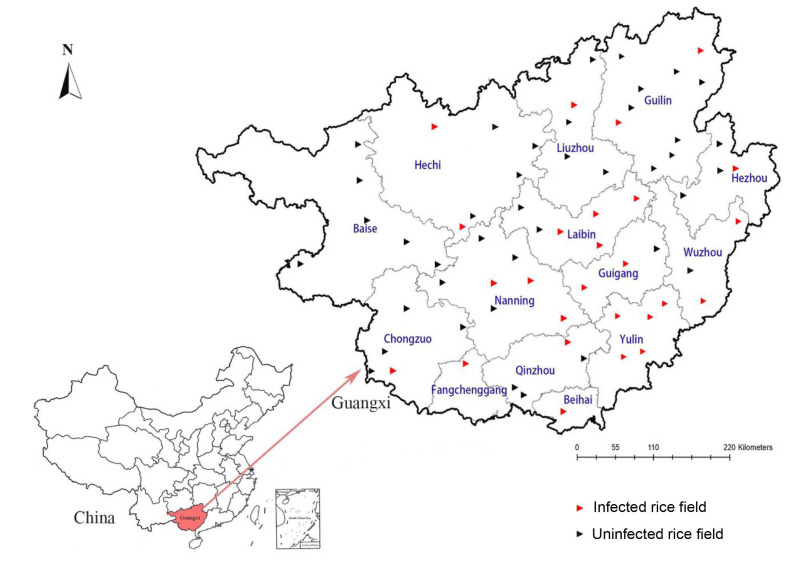
In China, M. graminicola was first discovered on Allium tistulosum in the Hainan province (Zhao et al., 2001). The nematode was then discovered on rice in China in Fujian, Hunan, Hubei, Zhejiang, Jiangxi and Sichuan Province (Song et al., 2017; Xie et al., 2019). Incidence of the disease is severe in the Hunan provinces, China. Disease incidence exceeded 85% in infected paddy fields (Song et al., 2017). In Guangxi, a survey of the RKN disease was performed during the rice-growing season of 2018-2019. Stunned, yellowish rice plants were found around 30 days after transplanting in fields. Galls and hooked tips on the roots and pear-shaped females within the galls were observed. The objectives of this study were to investigate the distribution of RKN on rice in Guangxi province and to identify species of RKN populations collected from different geographical localities in Guangxi using both traditional morphological features and molecular techniques.
In this study, a total of 206 fields was investigated during the rice-growing season in 67 counties of Guangxi. The RKN symptoms on rice were observed in 66 fields of 26 counties, and 32.04% fields were infested with RKNs (Table 1, Fig. 1). The females were collected by dissecting galls, and the second-stage juveniles (J2s) were obtained from the fresh rice roots with galls. The males were extracted from rhizosphere soil of rice using the method described by Liu (2000). Slides of nematodes were observed using a light microscope (Z2, the Carl Zeiss Axio Imager, Jena, Germany) for morphological characterizations. Measurements and pictures were taken using the ZEISS ZEN 2.6 (blue edition) software on a ZEISS Axiocam 506 microscope camera. Body length, maximum body width, dorsal pharyngeal gland orifice to stylet base, hyaline tail terminus, and stylet length were measured. The root tissues were teased apart with forceps and needles to remove adult females, and then perineal patterns were prepared and observed under a light microscope (Hunt and Handoo, 2009).
The perineal pattern of females (n = 9) showed dorsoventrally ovoid shape, with low to moderately high and round dorsal arches. Striae were smooth and continuous around the perineum, lateral lines were absent, and sometimes, a few lines converge at either end of the vulva, the character of this perineal pattern was good agreement with other description of M. graminicola reported, and was different from that of M. incognita and M. javanica with obvious lateral lines (Hunt and Handoo, 2009). Measurements (mean ± standard error, range) of males (n = 23) were obtained for body length (1,289.25 ± 149.49 μm, 1,040.26-1,581.30 μm), body width (43.33 ± 4.54 μm, 27.03-47.99 μm), stylet length (17.15 ± 0.76 μm, 15.29-18.41 μm), dorsal pharyngeal gland orifice to stylet base (3.61 ± 0.54 μm, 2.53-4.46 μm), and spicule length (29.81 ± 2.68 μm, 21.95-35.67 μm). Measurements of J2 (n = 21) included body length (470.80 ± 26.09 μm, 408.59-521.53 μm), body width (17.95 ± 1.55 μm, 15.73-22.29 μm), stylet length (12.57 ± 0.76 μm, 11.33-14.24 μm), and hyaline tail terminus (22.31 ± 2.54 μm, 17.82-26.07 μm) (Fig. 2). The morphology and morphometric values of males were matched well with the first description of M. graminicola reported by Golden and Birchfield (1965) and with those of other populations in China, except for body lengths, stylet lengths, and spicule lengths of this study are shorter than those of Hunan’s population (Table 2). The average body length of the J2s in our study was 470.80 μm, ranged from 408.59 to 521.53 μm, slightly longer than those of Sichuan, Zhejiang, and the first report. The J2s average stylet length of the Guangxi population is consistent with those of other Chinese populations, except for the Hunan population, which had slightly longer stylets. The hyaline tail terminus of Guangxi population was longer compared to other populations (Table 2). These small variations of morphometric are within the range of species variation of M. graminicola (Tian et al., 2018). As is known, the lengths of stylet, spicule, and vulva are considered as the key diagnostic parameters for the identification of M. graminicola. In previous studies, ten isolates of M. graminicola from Nepal, Bangladesh, USA, and India showed variations in morphometric measurements, while the internal transcribed spacer (ITS) sequences of all ten isolates were identified as M. graminicola (Pokharel et al., 2010). Similarly, the morphometric measurements of the Guangxi population also showed variation compare to different geographical populations from other provinces. It is possible that these morphometrical differences are due to geographical intraspecific variability and species variation, or phenotypic plasticity commonly exhibited by nematodes (Apolônio Silva de Oliveira et al., 2017; Pokharel et al., 2010; Salalia et al., 2017).
Ten J2s were handpicked and transferred individually to Eppendorf tubes with 19 μl of sterilized water for the confirmation of the morphological identification. DNA was extracted using the method of Qiu et al. (2016). Each nematode was frozen in liquid nitrogen for 1 min and heated in 85°C water bath for 2 min. The process repeated at least 3 times. The homogenate was incubated at 56°C in a polymerase chain reaction (PCR) buffer and 100 μg/ml proteinase K for 15 min and incubated at 95°C for 10 min. The ITS1-5.8S-ITS2 rDNA region was amplified with V5367(5′-TTGATTACGTCCCTGCCCTTT-3′) and 26S (5′-TTTCACTCGCCGTTACTAAGG-3′) (Vrain et al., 1992). The ITS region was amplified with species-specific primers Mg-F3 (5′-TTATCGCATCATTTTATTTG-3′) and Mg-R2 (5′-CGCTTTGTTAGAAAATGACCCT-3′) (Htay et al., 2016). The D2/D3 region of 28S RNA was amplified with D2A (5'-ACAAGTACCGTGAGGGAAAGTTG-3') and D3B (5'-TCGGAAGGAACCAGCTACTA-3') (De Ley et al., 1999). The PCR program of the thermal cycler consisted of the following thermal profile: 95°C for 5 min; 35 cycles of 30 s at 94°C, 52°C for 30 s, 72°C for 50 s; and a final extension cycle of 72°C for 10 min. Annealing temperatures were set up at 54°C for primers V5367/26S, 49°C for primers Mg-F3/Mg-R2, and 52°C for primers D2A/D3B. The amplified DNA fragments were excised from 1% agarose gel and purified using the DNA Purification Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions to purify the PCR products. The purified products were cloned into the pMD18-T vector system (TaKaRa Bio Inc., Kusatsu, Japan) and transformed into Escherichia coli strain DH5α chemically competent cells. Three representative colonies for each PCR product were selected, plasmid DNA carrying the insert was extracted and sequenced by using the pMD18-T Vector sequencing primers. Returned sequences were analyzed and edited manually using DNAMAN version 6.0. Consensus sequences obtained were compared with other nematode species available in the GenBank database through a BLASTn homology search program.
The ITS1-5.8S-ITS2 rDNA region yielded a PCR fragment of 771 bp. The sequences were deposited in GenBank as accession nos. MN0469602, MN647543, MN513027, and MN647593, had 87-100% similarity with those of M. graminicola isolation from Fujian, China (KM111531) and Pakistan (KX757064). The length of ITS region sequences of M. graminicola was 377 bp (Gen-Bank accession nos. MN128225, MN519726, MN511736, MN521459), and had 99-100% similarity with those of M. graminicola isolation from Sanya, China (MK613789), Vietnam (MG773553), and Brazil (MH703577). D2/D3 of 28S rDNA sequence of M. graminicola in this study was 773 bp, the region (GenBank accession nos. MN647592, MN513029, MN647589, MN647590, MN513028) was 99-100% identical to the sequences of M. graminicola USA isolation (AF435793 and AF435801) and Fujian, China isolation (HQ420904 and KM111529).
The phylogenetic trees based on aligned sequences of ITS1-5.8S-ITS2 rDNA, the D2/D3 of 28S rDNA and the ITS rDNA were constructed respectively by using the neighbor-joining method in MEGA5 software. M. graminicola populations formed a well-supported group within other species of Meloidogyne, and all Guangxi population (GenBank accession nos. MN049602, MN513028, MN519726, MN647543, MN513029, MN128225, MN513027, MN647592, MN511736, MN647593, MN647590, MN521459) appeared as a basal subclade in the M. graminicola clade, which is well-supported clade with other M. graminicola. Both morphological and molecular results confirmed that the nematode is M. graminicola.
Infectivity tests were confirmed in greenhouse conditions, twenty 2-week-old rice plants (cv. Teyou 09x10-3) were individually transplanted in pots with 1,000 cm3 autoclaved mixture of sand and field soil (sand:soil = 3:1). Each plant was inoculated 300 ± 35 J2s hatched from RKN samples eggs. Five rice plants non-inoculated were used as control. The roots were checked at 35 days after inoculation, obvious gall formations with typical hook-shaped root tips. No symptoms were found in control roots. Root galls were opened with a dissecting needle. Females and egg mass with eggs and J2s were observed. Morphological of perineal pattern and J2, and molecular characteristics of RKNs isolated from inoculated roots were identical to those in the field samples.
The disease caused by M. graminicola was severe in the suburb of Nanning city and Laibin city of Guangxi, China. To the best of our knowledge, this study presents the first report of M. graminicola on rice in Guangxi, China. Guangxi is an international terminal that links Southeast Asian Nations, an important area for double-crop rice culture with about 826.6 thousand hectares. This nematode is a severe problem to rice. Due to its ability to be transmitted by soil, agricultural machinery, and particularly by contaminated seeds, the nematodes pose a considerable threat to rice production in Guangxi, China. Further study will be conducted to screen resistant rice germplasm resources against the nematode and develop effective control strategies.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






