


In maintaining a consistent yield and quality of agricultural products, synthetic fungicides have played an important role through efficient control of plant diseases. However, safety and environmental problems caused by excessive use of synthetic fungicides instituted strict regulations on the registration of the active ingredients and application rates in fields (Paster and Bullerman, 1988). As an alternative to synthetic agents, microbial natural products have been considered as a source of diverse antifungal agents for plant disease control. Microbial natural products have been regarded as environment-friendly because their biodegradability could solve the residual problem of synthetic fungicides (Porter, 1985). Furthermore, owing to their versatile biological activity and structural diversity, microbial metabolites microbial metabolites are expected to provide new modes of action that would be effective against phytopathogenic fungi resistant to commercial fungicides (Kim and Hwang, 2007).
Among the fungal metabolites discovered so far, strobilurin A and B, isolated from the basidiomycete Strobilurus tenacellus, and several structurally-related compounds are the most famous fungal metabolites (Anke et al., 1977). Structural modification to improve the photostability and bioactivity of natural strobilurin enabled the commercialization of several strobilurin derivatives, such as azoxystrobin, pyraclostrobin, and kresoxim-methyl, as agricultural fungicides (Bartlett et al., 2002). Strobilurin binds to the ubihydroquinone oxidation center of the bc1 complex, which blocks electron transfer between cytochrome b and c1 in the mitochondrial membrane, thereby disturbing respiration and providing potent, broad-spectrum disease control efficacy at a parts-per-billion concentration (Bartlett et al., 2002; Clough et al., 1995; Kim and Hwang, 2007).
Trichoderma strains are one of the most studied fungal biocontrol agents and commercially available as biopesticides and biofertilizers (Harman et al., 2004). Various antibiotic compounds, including volatile and nonvolatile secondary metabolites, are produced by these strains. For instance, the volatile compound 6-pentyl-2H-pyran-2-one, exhibited in vivo and in vitro antifungal activity against plant fungal pathogens Rhizoctonia solani and Fusarium oxysporum f. sp. lycopersici (Scarselletti and Faull, 1994). Anthraquinone derivatives, including chrysophanol and pachybasin from Trichoderma harzianum, were also suggested to be involved in controlling R. solani (Liu et al., 2007).
Rhizopus soft rot disease caused by Rhizopus sp. is one of the most devastating postharvest diseases of various crops, including potato, stone fruits, and berries (Amadioha, 2001; Dennis, 1983; Qing and Shiping, 2000). Rhizopus species have a wide host range and grow fast in moist conditions (Salem et al., 2016). Rhizopus spores are spread into the air and mainly infect the wound sites of fruits and vegetables, followed by secrete strong cell wall-degrading enzymes to colonize the interior of host cells, resulting in serious economic losses during harvest, storage, and transport of fruits and vegetables (Qing and Shiping, 2000; Salem et al., 2016; Tang et al., 2012).
To reduce the loss in postharvest stages, various synthetic fungicides, such as iprodione, dichloran, fludioxonil, and tebuconazole, are applied (Bautista-Ba├▒os et al., 2014; Northover and Zhou, 2002). Due to residual toxicities and visible residues, however, only a few fungicides are approved for control of postharvest diseases (Zhang et al., 2020). Biological control of postharvest diseases of fruits by microbial antagonists has been considered as an alternative and effective strategy for reducing the postharvest loss of crops (Wisniewski and Wilson, 1992). Pichia, Cryptococcus, and numerous other yeast strains proved efficacious in controlling this fungus on stone fruit, strawberry, and tomato (Qing and Shiping, 2000; Roberts, 1990). Trichoderma harzianum has also been reported to control Rhizopus rot on apple and peach (Batta, 2007; El-Katatny and Emam, 2020). However, difficulties in maintaining a consistent application of the biological agent have increased interest in developing their antimicrobial metabolites as an alternative agent (Tripathi and Dubey, 2004; Usall et al., 2016). Pascale et al. (2017) studied the effect of harzianic acid and 6-pentyl-╬▒-pyrone isolated from T. harzianum and T. atroviride on grape (Vitis vinifera L.) infected with a powdery mildew fungus Uncinula necator. The foliar spray of these compounds suppressed the development of powdery mildew on grape leaves, as well as improved grape yield by 97% in terms of weight (Pascale et al., 2017).
In a filamentous fungal culture collection that we constructed, the fungal isolate F3736 was found to produce antifungal metabolites effective to inhibit the growth of Rhizopus oryzae in vitro. In the present study, the active compound was purified from the culture extract of F3736 through various chromatographic procedures and identified by spectroscopic analyses. The in vitro antifungal spectrum of the active compound and its control efficacy on Rhizopus rot of apple and peach fruits was also investigated.
Materials and Methods
Fungal material.
Setosphaeria rostrata F3736 was isolated from a wood debris collected in Gaeun Mountain, Seoul, Republic of Korea, in April 2016, and identified using the sequences of internal transcribed spacer region of rDNA by Korea University Culture Collection (KUC).
Purification of antifungal compound derived from S. rostrata F3736.
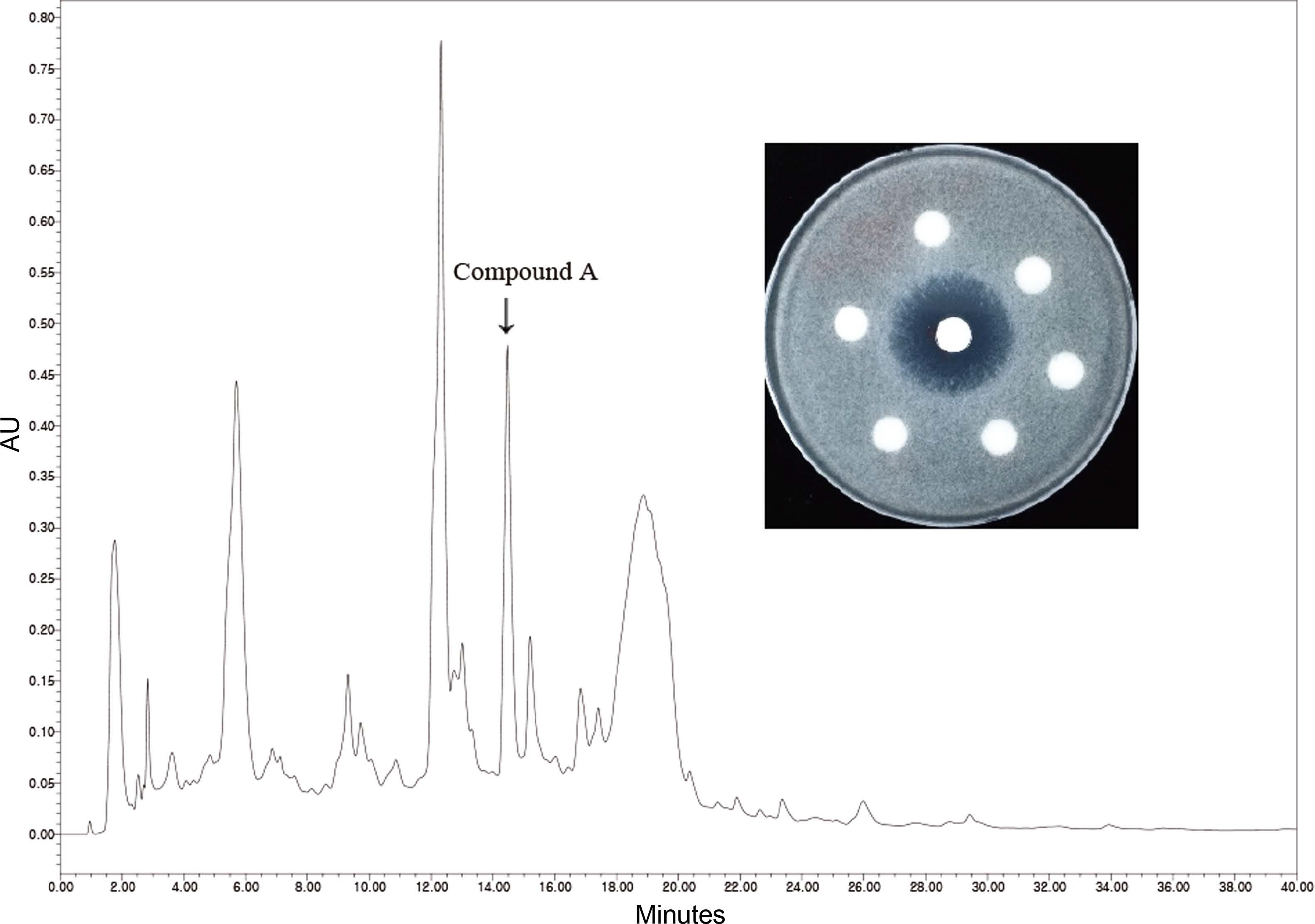
S. rostrata F3736 was cultured on potato dextrose agar (PDA; BD Difco, Franklin Lakes, NJ, USA) medium plates (3 l) at 25┬░C for 7 days and extracted with methanol. The methanol extract was filtered and concentrated to dryness, followed by re-suspended in 3 l of distilled water and re-extracted with same volume of ethyl acetate. The ethyl acetate layer was fractionated by Diaion gel column chromatography (Mitsubishi Chemical, Tokyo, Japan). The columns were eluted with a stepwise gradient of aqueous methanol (0%, 20%, 40%, 60%, 80%, and 100%, v/v) and washed by 500 ml acetone. The paper disk (8 mm in diameter; Advantec, Toyo, Japan) diffusion assay was used to assess the antifungal activity of each fraction against R. oryzae. The spore-seeded agar medium was generated by addition of 1 mL of conidial suspension (107 conidia/ml) of R. oryzae into 100 ml of molten PDA (0.7% agar). The paper disk absorbed with each fraction was placed on the spore-seeded agar medium and incubated at 28┬░C for 2 days. The active fractions were combined and further purified using a Varian HPLC system (Agilent Technologies, Santa Clara, CA, USA) equipped with a JŌĆ▓sphere ODS-H80 column (250 ├Ś 10 mm, 4 ╬╝m; YMC Co., Ltd., Kyoto, Japan). Chromatography was performed with a linear gradient of 20-95% aqueous methanol for 20 min and then isocratic elution (of 95% aqueous methanol) for 20 min at a flow rate of 1 ml/min. The eluates were monitored at an absorbance of 230 nm. The fractions were manually collected peak by peak and bioassayed against R. oryzae. The antifungal fraction was denoted as compound A.
Structural elucidation of purified bioactive compound.
The molecular weight of compound A was determined by electrospray ionization mass spectrometry using an ultraperformance liquid chromatography-quadrupole time-of-flight mass spectrometer at the Seoul Branch of the Korean Basic Science Institute. The nuclear magnetic resonance (NMR) spectra of compound A was recorded using a Varian 500-MHz nuclear magnetic resonance spectrometer (Varian, Palo Alto, CA, USA) equipped with a 5-mm AutoX DB probe. The 1H- and 13C-NMR data along with 1H-1H correlation spectroscopy, 1H-13C heteronuclear multiple bond correlation, and 1H-13C heteronuclear single quantum correlation spectra of compound A measured in chloroform-d (Cambridge Isotope Laboratory, Andover, MA, USA). Chemical shifts were referenced to solvent peaks (╬┤H 7.26 ppm and ╬┤C 77.0 ppm). Based on the spectroscopic data, the structure of compound A was determined.
Effect of moriniafungin on various plant fungal pathogens.
The minimum inhibitory concentration (MIC) values of purified moriniafungin were evaluated against various plant pathogenic fungi using the modified CLSI M38-A method (Espinel-Ingroff et al., 2005). The 25 ╬╝l of conidial suspension (4 ├Ś 105 conidia/ml) of A. brassicicola, A. oryzae, B. cinerea, C. gloeosporioides, C. orbiculare, F. oxysporum f. sp. lycopersici, R. oryzae, or R. stolenifer var. stolonifer, together with 25 ╬╝l of quadruple-strength potato dextrose broth (PDB; BD Difco) were added to 96-well plates (SPL life Sciences, Pocheon, Korea). Subsequently, moriniafungin was added to each well at final concentrations of 0.01, 0.02, 0.03, 0.06, 0.13, 0.25, 0.50, 1, 2, 4, 8, 16, 32, and 64 ╬╝g/ml. After 28┬║C incubation for 2 days, MICs of compound A against test fungi were determined by visual observation.
In vivo assessment of Rhizopus rot control efficacies.
The protective control efficacies of moriniafungin and tebuconazole (as a positive control) against Rhizopus rot were assessed on peach and apple fruits. Purified moriniafungin and a commercial fungicide tebuconazole were prepared in water containing 0.05% Tween-20 (v/v) and serially diluted to 0.04, 0.20, 1, 5, and 25 ╬╝g/ml. Fruits were purchased from a local market for the experiment. The conidia (1 ├Ś 106 conidia/ml) of R. stolenifer var. stolenifer and R. oryzae were harvested with PDB containing 0.05% Tween-20 after cultivation for 1 week on PDA. Peach and apple fruits were surface disinfested in 1% sodium hypochlorite, washed, and wounded (three wounds per fruit) using a sterile blue tip (Axygen Scientific, Inc., Union City, CA, USA). Each solution (5 ╬╝l) of moriniafungin and tebuconazole were applied to the wound site of peach and apple fruits. After air drying for 1 h, conidial suspensions (5 ╬╝l) of R. stolonifer var. stolonifer and R. oryzae were inoculated onto wound sites of peach and apple fruits, respectively. The fruits were placed in a humidified plastic container kept at 28┬║C. Four days after inoculation, the lesion diameter (mm) of Rhizopus rot in each fruit was measured. Each treatment was performed in three replicates.
Results and Discussion
Purification of antifungal compound produced by Setosphaeria rostrate.
In this study, the culture extract of S. rostrata strain F3736 showed inhibitory activity against R. oryzae. The antifungal compound was isolated from the culture extract and was identified as moriniafungin. The methanolic extract of S. rostrata was partitioned with ethyl acetate and purified by a series of chromatography steps. The Diaion HP-20 column loaded with the ethyl acetate extract fraction produced 80% and 100% MeOH fractions, which were bioactive against R. oryzae. The active fractions were further purified by reverse-phase highperformance liquid chromatography (HPLC) with detection at 230 nm. The peak (denoted as compound A) eluted at the retention time of 14.5 min showed antifungal activity against R. oryzae (Fig. 1). The active peak eluate was collected through a repeated preparative HPLC procedure and lyophilized to white powder (3.5 mg).
Structure elucidation of moriniafungin.
Electrospray ionization mass spectra of compound A recorded m/z 689.3537, [M-H]- (calculated, 689.3540 as C37H53O12) in negative mode, m/z 713.3508, [M+Na]+ (calculated, 713.3513 as C37H54NaO12) in positive mode (Fig. 2). Based on the MS and NMR experimental data (Fig. 2, Supplementary Figs. 1-5) in comparison with literature data (Basilio et al., 2006; Zhang et al., 2019), the structure of compound A was identified as moriniafungin (Fig. 3). The 1H- and 13C-NMR data of compound A were summarized in Table 1.
Moriniafungin is a derivative of sodarin (an antifungal metabolite possessing a tetracyclic diterpene glycoside structure) that inhibits fungal protein synthesis (Dom├Łnguez et al., 1998). Sordarin and their derivatives were originally found by screening protein synthesis inhibitors using a cellfree translational system (Basilio et al., 2006). The mode of action of sordarin was shown to be stabilization of the elongation factor 2 (EF2)-GDP-ribosome complex in fungi (Dom├Łnguez et al., 1998; Justice et al., 1998). The specific binding of sordarin to fungal EF2 rather than the EF2 of plants and mammals suggested that sordarin and its derivatives have potential as antifungal drugs with selective toxicity against fungal pathogens in mammalian and plant hosts (Deshmukh et al., 2016).
Moriniafungin contains a 2-hydroxysebacic acid residue linked to the C-3ŌĆ▓ of the sordarose moiety through a 1,3-dioxolan-4-one ring. Basilio et al. (2006) suggested this kind of modification at C-3ŌĆ▓ in the sordarose residue can increase the antifungal activity and also broaden the antifungal spectrum compared with sordarin, which could also be applied to other sordarin derivatives, like zofimarin (Basilio et al., 2006; Daferner et al., 1999; Tanaka et al., 2002). Additionally, C-3ŌĆ▓ sordarin analogs possessing a side chain with a terminal carboxylic acid are suggested to be involved in binding to EF2, an action thought to be partly responsible for increasing the antifungal spectrum and potency of moriniafungin compared with other C-3ŌĆ▓ sordarin analogs (Deshmukh et al., 2016).
In vitro antifungal activities of moriniafungin.
The MIC values of moriniafungin against phytopathogenic fungi were summarized in Table 2. Among test phytopathogenic fungi, the genus of Colletotrichum and Rhizopus causing anthracnose and soft rot diseases on various fruits and vegetables was most sensitive to moriniafungin. Notably, moriniafungin was effective to inhibit the growth of R. oryzae and R. stolenifer var. stolenifer at very low concentrations (0.03-0.13 ╬╝g/ml). At higher concentration (1-8 ╬╝g/ml), the growth of C. gloeosporioides and C. orbiculare was also completely inhibited by moriniafungin. However, there was no inhibitory effect of moriniafungin against Alternaria, Aspergillus, Botrytis, and Fusarium spp. at the highest concentration tested (64 ╬╝g/ml).
In determining the MIC values of moriniafungin against phytopathogenic fungi, moriniafungin showed the selective antifungal activity against R. oryzae and R. stolenifer var. stolenifer with the lowest MIC values of 0.0313 and 0.125 ╬╝g/ml, respectively. Similar results have been reported in previous studies on sordarin derivatives. Hydroxysordarin displayed the lowest MIC value of 1 ╬╝g/ml against a zygomycete fungus Mucor miehei, while could not inhibited the growth of other filamentous fungi including Alternaria sp. B. cinerea, and Fusarium spp. (Daferner et al., 1999). Similarly, sordarin and GM237354 (a semi-synthetic derivative of sordarin) showed the narrow inhibitory activity against zygomycete fungi including species Absidia, Cunninghamella, Mucor and Zygorhynchus (Daferner et al., 1999; Herreros et al., 1998). Similar to the literature data, in our study, the sordarin analog moriniafungin displayed a narrow antifungal spectrum, with modest antifungal activity against the ascomycetes Colletotrichum spp. but potent activity against the zygomycete fungi Rhizopus spp.
Protective control efficacies of moriniafungin against Rhizopus rot.
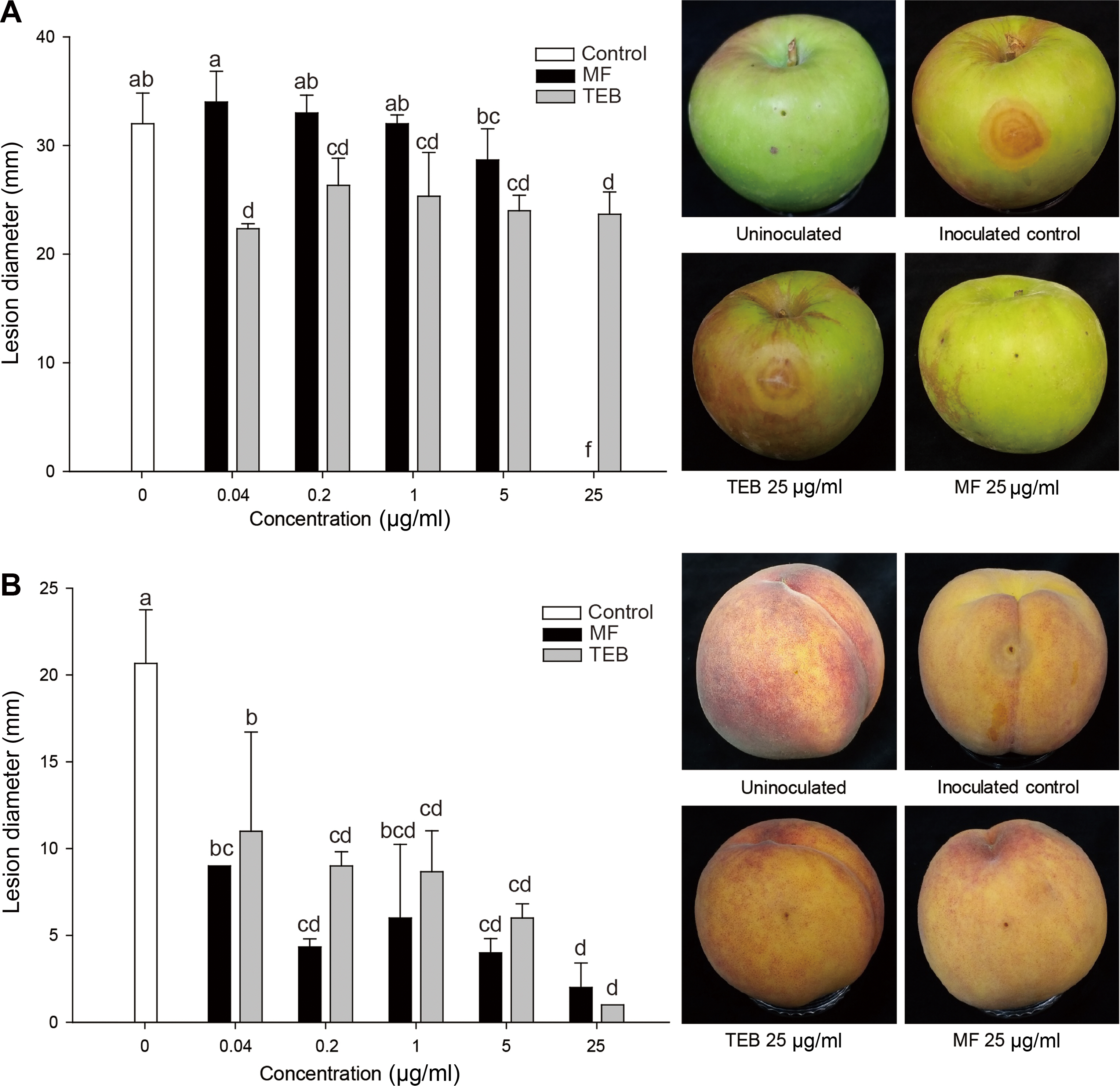
With the consideration that moriniafungin showed significant inhibitory effects on Rhizopus spp. in the microdilution assay, we examined the disease control efficacies of moriniafungin on Rhizopus rot of apple and peach fruits. Four days after inoculation of R. oryzae, the typical soft rot symptoms including brownish discoloration, softening, and exudation from wounds were observed on the non-treated apple fruits (Fig. 4A). The mean diameter of the lesions of the inoculated control was 32.00 ┬▒ 2.83 mm. Moriniafungin (0.04-5 ╬╝g/ml) showed no significant effect on reducing apple rot lesions caused by R. oryzae (Fig. 4A). However, at higher concentration (25 ╬╝g/ml), moriniafungin completely protected apple fruits from the attack of R. oryzae (no symptom was observed around the inoculated area). The disease control efficacy of morin iafungin (25 ╬╝g/ml) was superior to that of a synthetic fungicide tebuconazole. Even at the highest concentration of tebuconazole (25 ╬╝g/ml), apple fruits were discolored, softened, and water-soaked by R. oryzae (Fig. 4A).
Furthermore, moriniafungin effectively controlled the Rhizopus rot development of peach fruits caused by R. stolonifer var. stolonifer (Fig. 4B). The mean diameter of lesions on the inoculated control was 20.67 ┬▒ 3.09 mm. At all applied concentrations, moriniafungin and tebuconazole reduced rot disease development significantly compared with the non-treated control. At concentrations of 0.04-5 ╬╝g/ml, moriniafungin significantly reduced the lesion diameter by 56.5-80%, compared with the non-treated control. When treated with moriniafungin (25 ╬╝g/ml), more than 90% reduction in diameter of lesions was observed. The inhibitory activity of moriniafungin against infection and colonization of R. stolenifer var. stolenifer on peach fruits was comparable to that of a synthetic fungicide tebuconazole (Fig. 4B).
Considering the potent inhibitory activity of moriniafungin against Rhizopus spp. in vitro, moriniafungin was examined for controlling Rhizopus rot of apple and peach fruits and thus evidently reduced the development of discolored, softened, and water-soaked lesions on fruits (Fig. 4). The in vivo antifungal effect of moriniafungin was comparable to that of tebuconazole, an effective fungicide to control postharvest Rhizopus rot of wounded peach fruits (Northover and Zhou, 2002).
For several decades, fungal metabolites have been investigated as control agents of postharvest diseases. 6-Pentyl-2-pyrone produced by Trichoderma spp. was reported to be effective in reducing the incidence of B. cinerea storage rots in kiwifruit (Bonnarme et al., 1997; Poole et al., 1998). 6-Pentyl-2-pyrone is a naturally-occurring compound also found in ripening peach and nectarine, which makes it acceptable as a food flavoring additive or postharvest treatment (Horvat et al., 1990). Fusapyrone and deoxyfusapyrone, identified from the culture of Fusarium semitectum, showed inhibitory activity against B. cinerea on grapes at 100 ╬╝g/ml and displayed no phytotoxicity and low toxicity in mammals, which makes them potential agents for postharvest disease control (Altomare et al., 2000).
This study revealed that moriniafungin, isolated from the culture extract of S. rostrata strain F3736, could inhibit the growth of several species of plant pathogenic fungi. Especially, the MIC of moriniafungin against Rhizopus spp. was an order of magnitude lower than that of tebuconazole, the commercial fungicide used as the positive control. Moriniafungin reduced the lesion development on apple and peach fruits significantly compared with the non-treated control, which was comparable to the treatment of tebuconazole at the same concentrations. In this study, the sordarin derivative moriniafungin, as a specific inhibitor of fungal EF2, was suggested as a potential control agent of postharvest diseases caused by Rhizopus species.
Electronic Supplementary Material
Supplementary materials are available at The Plant Pathology Journal website (http://www.ppjonline.org/).


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






