Pathogenicity Determinants of Sclerotinia sclerotiorum and Their Association to Its Aggressiveness on Brassica juncea
Article information

Abstract
White rot or stem rot caused by Sclerotinia sclerotiorum is one of the most destructive fungal diseases that have become a serious threat to the successful cultivation of oilseed Brassicas. The study was designed with an aim to investigate the association between the pathogenic aggressiveness and pathogenicity determinants of this pathogen specifically in Brassica for the first time. For this, a total of 58 isolates of S. sclerotiorum from different geographical regions were collected and purified. These isolates were inoculated on a Brassica juncea cv. RL-1359 and they exhibited high level of variation in their disease progression. The isolates were grouped and then 24 isolates were selected for the biochemical analysis of pathogenicity determinants. The isolates varied significantly with respect to their total organic acids, oxalic acid production and pectin methyl esterase and polygalacturonase activity. The oxalic acid production corresponded to the disease progression of the isolates; the isolates with higher oxalic acid production were the more aggressive ones and vice-versa. This is, in our knowledge, the first study to establish a correlation between oxalic acid production and pathogenic aggressiveness of S. sclerotiorum on B. juncea. However, the pectinases’ enzyme activity did not follow the trend as of disease progression. These suggest an indispensable role of oxalic acid in pathogenicity of the fungus and the potential to be used as biochemical marker for preliminary assessment of pathogenic aggressiveness of various isolates before incorporating them in a breeding program.
Sclerotinia sclerotiorum (Lib.) de Bary, the causal of stem rot or white mould, is a non-host specific, necrotrophic soil-borne pathogen that attacks a wide range of cultivated and wild species including rapeseed-mustard. The fungus infects over 400 species of plants worldwide (Boland and Hall, 1994). Earlier, this disease was considered to be of little economic importance in rapeseed-mustard, but in recent years it has gained the importance of being the most wide-spread and destructive disease. In this crop, the disease manifests mainly as stem rot, resulting in crop lodging and severe losses. Yadav et al. (2011) reported yield losses up to 40%, while in severe incidence, losses may go up to 74% (Sharma et al., 2001b).
The fungus produces hard resting structures; the sclerotia, that remain viable in the soil for many years. On imbibation of water from soil, sclerotia germinate either myceliogenically (produce mycelia) or carpogenically (produce apothecia) depending upon the environmental conditions (Steadman, 1983). Upon infection on stem, initially water-soaked lesions are formed which expand rapidly and are covered with whitish mycelial growth (Bolton et al., 2006). Mycelial infection in the field is generally very patchy, whereas ascosporic infection is more uniform. When the stem is completely girdled and covered by the whitish mycelial growth all over, the plant wilts and dries. The fungus produces symptoms on all the aerial plant parts but the most destructive ones are produced on the stem. Later in the season, sclerotia are formed inside the pith of the stem as well as on the outside mycelial growth on the infected tissue. Usually the plants are attacked at flowering stage and once the pathogen is established in the field, it is very difficult to eliminate it due to the stubborn soil-borne sclerotia and wide host range.
Though, cultural practices like crop rotation with grasses and management of irrigation along with the use of some fungicides, offers some reduction in disease severity, but effective management of this fungal disease is still lacking (Ando et al., 2007). The most sustainable and efficient way to manage this disease is host plant resistance (Li et al., 2006); thus, the first important step is identification of reliable sources of resistance and there-after incorporation in the elite cultivars. At present, there are no known sources of complete and robust resistance against it. However, partial resistance has been identified by various workers in mostly B. napus and very few in B. juncea (Li et al., 2006, 2008; Zhao et al., 2004). Identifying sources of resistance in Brassica against Sclerotinia is challenging due to the variability existing in the aggressiveness of the pathogen population. Thus, for effective screening of germplasm and identification of reliable resistance sources, it is very important to have an insight of the variability in the population. This has focused our attention on the need of detailed study and understanding of the pathogenic factors involved in disease development.
S. sclerotiorum is responsible for secreting a wide array of substances to facilitate its necrotrophic life style (Annis and Goodwin, 1997). Oxalic acid seems to be an indispensable pathogenicity determinant (Zhou and Boland, 1999). Corroboration of this involvement of oxalic acid is based upon the oxalate recovery from infected tissues in millimolar concentrations (Marciano et al., 1983). The positive correlation between oxalic acid and pathogenicity has also been strengthened by the study, where mutants of Sclerotinia lacking in the ability to synthesize oxalate were nonpathogenic, whereas, the revertant strains after regaining their oxalate biosynthetic ability, exhibited normal virulence (Godoy et al., 1990). The fungus primarily utilizes the mechanical pressure to penetrate the host cuticle; however, lytic enzymes are also known to play an important role in it (Saharan and Mehta, 2008). Tissue maceration, cell wall lysis, and solubilization of cell wall constituents is brought about by strong and repeated action of various lytic enzymes (Cessna et al., 2000). In this instance, cell wall degrading enzymes (CWDEs) including predominantly pectinases along with cellulases, hemicellulases, xylanases, glucanases etc. act and bring about maceration of the host tissue, followed by necrosis and ultimately death of the plant (Riou et al., 1991). Amongst pectinolytic enzymes, S. sclerotiorum is known to produce various isoforms of exo- and endo-polygalacturonases (PGs) and pectin methyl esterases (PME) (Waksman et al., 1991). The PME deesterifies the methylated pectin producing demethylated pectin, which is then acted upon by PG enzyme, breaking it down to galacturonic acid units, thus solubilizing the pectin as whole.
Keeping in view the above-mentioned, this study was designed (1) to evaluate the production of various pathogenicity determinants by different S. sclerotiorum isolates and (2) to establish an association between these and pathogenic aggressiveness of the isolates.
Materials and Methods
Evaluation of pathogenic aggressiveness of S. sclerotiorum isolates
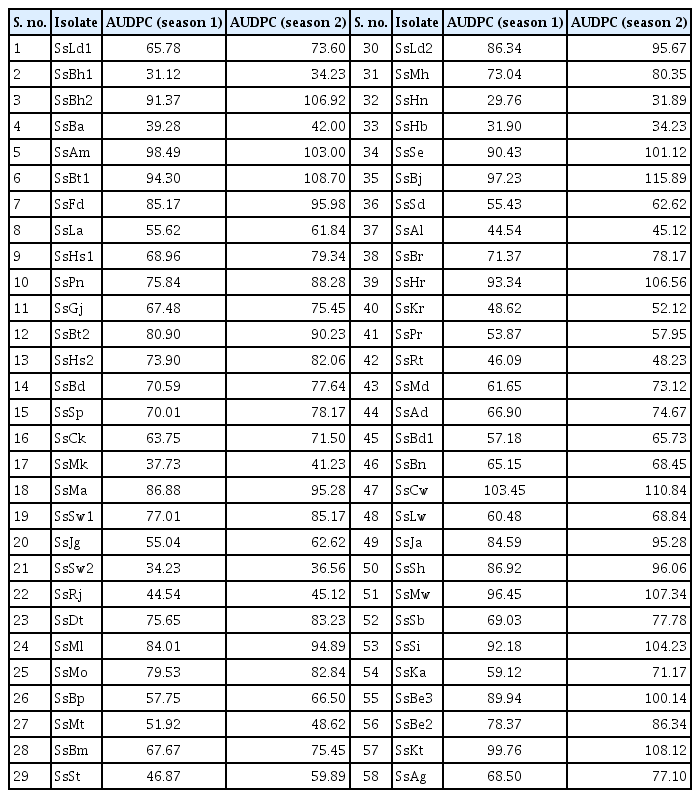
In this study, a total of 58 isolates of S. sclerotiorum, collected from various rapeseed-mustard growing areas of Punjab and adjoining states were used (Table 1). B. juncea cv. RL 1359 was used to evaluate all the isolates of S. sclerotiorum for their pathogenic aggressiveness for two years. Plants were inoculated using standard stem inoculation technique (Buchwaldt et al., 2005). The inoculated plants were given frequent irrigations and sprinklers were used at regular intervals to maintain optimum humidity. Data was recorded as lesion length at 1, 2, and 3 weeks after inoculation and area under disease progression curve (AUDPC) was worked out for each isolate.

Isolates of Sclerotinia sclerotiorum and their place of collection
Total acids production
Total acids production was estimated by the procedure described by Durman et al. (2005). The isolates were grown on bromophenol blue (BPB) amended potato dextrose agar medium and incubated for 3 days in darkness at 22 ± 2°C. Observation on diameter of mycelial growth in the colony (C) and yellow color developed (Y) were measured and the ratio (Y/C) was used as a measure to compare different isolates for total acids production.
Oxalic acid and pectinase enzyme analysis
As followed by Sharma et al. (2001a), the production of oxalic acid, PG, and PME was standardized w.r.t. growth period of S. sclerotiorum. For this, a single isolate of S. sclerotiorum (SsAm) was grown and filtrate was extracted every day up to 10 days of incubation for determination of oxalic acid content and pectinases activity.
For oxalic acid determination, the filtrate was obtained by growing isolates in potato dextrose broth for 3 days. The filtrate was centrifuged and the supernatant was analyzed using the procedure described by Xu and Zhang (2000). The mycelium dry weight was recorded after keeping it at 80°C for 72 h. For enzymatic analysis, the isolates were grown in Czapek dox’s broth with 1% pectin as sole carbon source for 7 days. The activity of PME and PG was estimated by the methods described by Desse and Stahmann (1962) and Miller (1959), respectively.
Statistical analysis
All assays were run in triplicate and data was subjected to analysis of variance (ANOVA) and compared using the Duncan grouping test at 5% probability (P < 0.05), with the assistance of the SAS program (SAS Institute Inc., Cary, NC, USA).
Results
Aggressiveness studies of S. sclerotiorum isolates
The isolates of S. sclerotiorum differed considerably w.r.t. the disease progression on B. juncea cv. RL 1359 (Table 2, Fig. 1). The AUDPC values across the isolates in both seasons exhibited a range from 29.76 (SsHn) to 115.89 (SsBj). The isolates were categorized into five different groups based upon their AUDPC values (Table 3).
Area under disease progression curve for Sclerotinia sclerotiorum isolates on Brassica juncea cv. RL 1359 (season 1 & 2)

Initial symptoms (A) and various kinds on lesions (B-H) formed by different isolates on Brassica juncea cv. RL-1359.

Grouping of isolates according to their AUDPC values on Brassica juncea cv. RL-1359
Analysis of pathogenicity determinants
For this biochemical analysis, 24 representative isolates were selected from different groups based on the AUDPC values and were further used (Table 3).
Standardization
It was observed that oxalic acid production increased up to 3 days of incubation while pectinases’ activity increased up to 7 days of incubation and declined as growth further progressed (Fig. 2). Therefore, for analysis of all the isolates, filtrate was obtained at 3 days and 7 days of incubation for oxalic acid and pectinases, respectively.

Oxalic acid production (A), polygalacturonase (PG) (B), and pectin methyl esterase (PME) enzyme activity (C) with respect to growth of Sclerotinia sclerotiorum.
Total acids production
As the fungal mycelium grew, it produced various acids which diffused through the media and changed the colour of BPB from violet to yellow (Fig. 3). The Y/C ratio served as the measure of total acids production and the isolates exhibited a substantial amount of variability in total acids production with the isolate SsHr and SsBe2 having the highest Y/C ratio (0.94) and isolate SsBa having the lowest one (0.44).

Yellow colour developed due to acid production by growing mycelium of Sclerotinia sclerotiorum in bromophenol blue amended potato dextrose agar medium.
Oxalic acid analysis
The results revealed that the mean oxalic acid production ranged between 927.9 (SsBa) and 6,060.5 (SsBe2) μg oxalic acid/g dried mycelium. Thus, a significant variation was observed among the isolates w.r.t. oxalic acid production (Table 4). A high positive correlation was found between oxalic acid and total acids production (r2 = 0.9). On comparing the trend of oxalic acid production to that of AUDPC of these isolates (Fig. 4), it was observed that overall trend of oxalic acid coordinated with that of AUDPC. Except for four isolates, SsRt, SsSd, SsLw, and SsSb, all the isolates were having oxalic acid production complimenting their AUDPC value (r2 = 0.7).

Ratio of diameter of the yellow colour developed (Y) and diameter of the colony (C) for total acids production

Relation of oxalic acid production (A), polygalaturonase (B) and pectin methyl esterase activity (C) and area under disease progression curve (AUDPC) of Sclerotinia sclerotiorum isolates.
PG and PME enzyme analysis
All the isolates varied with respect to the enzyme activity (Table 4). The highest PG enzyme activity was observed in culture filtrate of isolate SsSd (9.44 μmoles of reducing sugar released/ml enzyme/min) while the lowest (0.41 μmoles of reducing sugar released/ml enzyme/min) was observed for isolate SsSw2. In between these, the isolates SsSw2, SsBh1 and SsMh exhibited lower activity and SsAm, SsSd, SsBr, SsHr, SsLw, and SsBe2 exhibited higher enzyme activity. The highest PME activity was observed in the isolate SsSd (42.56 methoxyl groups liberated in μ equivalents/min/ml) and the lowest was observed in isolate SsBd (26.44 methoxyl groups liberated in μ equivalents/min/ml). Activities of all the other isolates ranged in between with SsBh2 and SsBa exhibiting activity (41.22 methoxyl groups liberated in μ equivalents/min/ml) close to highest value and SsMl, SsAl, SsLw, and SsSh exhibiting activity (27.78 methoxyl groups liberated in μ equivalents/min/ml) near to the lowest value.
Unlike oxalic acid, overall trend of the activity of both the pectinases did not coordinated well with that of the AUDPC values of the isolates (Fig. 4). Correlation coefficient values of PG and PME with the AUDPC values were 0.4 and zero, respectively. The enzyme activity was found to be indicating no robust correlation with pathogenic aggressiveness. Isolates SsBh1, SsSw2, SsBa, and SsRj had lower AUDPC values and their PG activity was also on the lower side, SsBm, SsSp, and SsBp are in the mid zone of AUDPC and PG enzyme activity values and SsBe2, SsHr, and SsAm had higher AUDPC values and their enzyme activity is also higher. Except for the mentioned isolates, for other isolates, AUDPC values did not correspond to their enzyme activity values. Isolate SsHr having second highest AUDPC value and isolate SsSw2 having second lowest AUDPC possessed same activity of PME.
Discussion
The results gave us a deeper and detailed knowledge of pathogenic determinants produced by S. sclerotiorum. As 58 isolates were analyzed in this study; the variation present in the lesion lengths and disease progression of different isolates on same cultivar, under same conditions, point towards the underlying variation in the pathogenic aggressiveness of these isolates. For resistance breeding to be a success, these underlying differences within the pathogen population are important to be known so that stable and reliable sources of resistance can be identified and selected to be carried forward in these programs.
During compatible interaction with its host, S. sclerotiorum secretes a full array of CWDEs to facilitate its penetration, carry out tissue maceration and degradation of various cell-wall components (Riou et al., 1991). Pectinases secreted by S. sclerotiorum play major role in lysis and degradation as pectin is the major component of plant cell wall and middle lamella. In addition to these enzymes, the toxin secreted by this fungus: oxalic acid, is known to play a pivotal role in disease development and thus is a key pathogenicity determinant (Lumsden, 1979). Besides oxalic acid, is also known to release other dicarboxylic organic acids such as succinic, malic, fumaric, and glycolic acids (Vega et al., 1970). In this study, oxalic acid production and disease progression (as AUDPC) were highly positively correlated. The isolates on higher side of the AUDPC values were also on the higher side of oxalic acid production and vice-versa for lower side. This suggests that pathogenic aggressiveness of S. sclerotiorum has a high dependency on oxalic acid production. To our knowledge, this is the first study to have correlated oxalic acid production in S. sclerotiorum isolates to its pathogenic aggressiveness on Brassica juncea. The evidence for the involvement and such dependence of pathogenicity on oxalic acid is based on its recovery from infected tissues (De Bary, 1866; Marciano et al., 1983) and the correlation between disease score/severity and oxalic acid production in some previous studies. Durman et al. (2005) reported significant differences amongst different mycelial compatibility groups’ w.r.t. their oxalic acid and other organic acids production. S. sclerotiorum oxalic acid deficient mutants were non-pathogenic even when they were having their full battery of CWDEs on dry beans whereas their revertant strains, that regained their ability to produce oxalic acid displayed normal virulence patterns (Godoy et al., 1990). The mechanism(s) by which oxalic acid aids in pathogenicity are; it provides optimum conditions for CWDEs to work by lowing pH to 4–5, helps in escaping the attack of polygalaturonase inhibiting proteins (Favaron et al., 2004), chelates Ca2+ and makes way for PG to hydrolyse the pectates and suppresses the oxidative burst in host plants (Guimaraes and Stotz, 2004). This explains the correlation and the similar trend obtained between the aggressiveness and oxalic acid secreted by the isolates in this study.
De Bary (1866) was the first to suggest that extracellular enzymes may be involved in the infection process of plant pathogenic fungi. Of the numerous CWDEs produced by plant pathogenic fungi, most research has focused on the pectin degrading enzymes. This is because the pectinases are typically produced first, in the largest amounts and are the only CWDEs capable of macerating plant tissue and killing plant cells on their own. PME carries out the demethylation of esterified pectin and thus makes it available for other enzymes, like polygalaturonase, for further break down to galacturonic acid monomers. The isolates used in this study varied considerably in pectinase enzyme activity but a correlation could not be worked out with AUDPC of the isolates. In addition to these, many other CWDEs are produced by this fungus. This may be attributed to the fact that pathogenic aggressiveness is not governed by these enzymes only but is determined by various other factors involved along with these pectinases. Many other CWDEs (cellulases, hemicellulases, xylanases, glucanases, proteases, etc.) are produced by this fungus; thus, these particularly studied enzymes won’t be acting and governing the pathogenicity just on their own. The expression of most CWDEs is tightly regulated at the transcription level by the availability of carbon and/or nitrogen sources and also the pH levels (Rollins and Dickman, 2001). A study in Saskatchewan was conducted using 114 isolates of S. sclerotiorum for their disease causing capacity and pectinase enzyme production (Morrall et al., 1972, 1976) and concluded that an endo-PG was the most prevalent of the entire pectic enzymes but there was no significant positive correlation between isolate pathogenicity and its pectinolytic enzyme activities. This also suggests that any one pectolytic enzyme might not be rendered soleley responsible for the pathogenicity and a programmed play of lytic enzymes is there which is governing the pathogenicity. During interaction with the host, the monomers released from host tissues degradation brought about by pathogenic origin lytic enzyme action and the presence of other pathogenicity determinants might be acting as inducers for stimulating more production of pectinases and thus we were not able to get that in-vivo.
This study gave us a clear insight of difference existing within the isolates of S. sclerotiorum with respect to their pathogenic aggressiveness, which could be very well correlated to their oxalic acid production but not so to the pectinase enzyme activity. Thus, the oxalic acid production can be used as a biochemical marker to study different isolates and their pathogenic aggressiveness may be anticipated. This will ensure involvement of aggressive isolates of S. sclerotiorum in the screening programs and thus can be of real help to identify robust resistance sources. The assay being simple and easy to carry out will surely help breeders and pathologists to identify aggressive isolates before taking the things to field testing, which will save time and other resources. As far as pectinases’ activity is concerned, further studies may be required to carry out for analyzing the whole enzymatic profile during the host-pathogen interaction so that it come out more clearly which enzymes or enzyme combinations have more effect on governing the pathogenicity.
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.