



Social pressure on caring for the environment and the concept of One Health is increasingly prevalent at the level of consumers and society as a whole (Andrivon et al., 2022). In this sense, the genetic control of diseases together with the increase in yield has been prioritized by breeders (Brown, 2015). Durable resistance to bacterial and fungal diseases continues to be a challenge in breeding programs. Bacterial wilt is a widespread vascular disease that affects more than 450 plant species in tropical, subtropical, and temperate regions of the world (Elphinstone, 2005), some of great economic importance such as potato, tomato, bell pepper, eggplant and tobacco (Hayward, 1991). Ralstonia solanacearum responsible for this disease, is a soil-borne pathogen that enters the plant through wounds in the root tissues and progressively invades the vascular tissues, causing partial or total wilting and, finally, the death of the plant (Hayward, 1991). Vascular diseases have been shown to be the most destructive to annual crops and woody perennials. Because vascular pathogens live inside their host plants, they make cultural, chemical and biological control measures inefficient, highlighting the need to design high-level strategies to combat them (Yadeta and Thomma, 2013). Given the complexity of its control under production conditions, host resistance is the most sustainable and environmentally effective strategy. However, there are currently no potato cultivars resistant to R. solanacearum available for production.
The incorporation of resistance by genetic engineering has generated fast and safe alternatives to traditional breeding (Dong and Ronald, 2019, 2021). An interesting strategy is the expression of pattern recognition receptors (PRRs) to enhance the immune response, and thus generate resistance to pathogens (Boller and Felix, 2009; Gurr and Rushton, 2005; van Esse et al., 2019). Plants recognize the presence of pathogens through PRRs located on the cell surface and intracellular nucleotide-binding site leucine-rich repeat receptors (Lu and Tsuda, 2021). Plant PRRs recognize molecular patterns associated with pathogens or cell damage (PAMP or DAMP, respectively) leading to activation of immunity (PAMP-triggered immunity). In particular, the Arabidopsis thaliana immune EF-TU RECEPTOR (EFR) recognizes bacterial elongation factor Tu, triggering an immune response. The EFR gene transfer has been shown to increase resistance in Nicotiana benthamiana against Agrobacterium tumefaciens and Pseudomonas syringae pv. tabaci, in tomato against R. solanacearum (Lacombe et al., 2010), in rice against Xanthomonas oryzae pv. oryzae (Lu et al., 2015; Schwessinger et al., 2015), in wheat against Pseudomonas syringae pv. oryzae (Schoonbeek et al., 2015), in orange against Xanthomonas citri subsp. citri (Xcc) and Xylella fastidiosa subsp. pauca (Xfp) (Mitre et al., 2021), and in Medicago truncatula against R. solanacearum (Pfeilmeier et al., 2019; Piazza et al., 2021).
Recently, our group characterized the introgression of the EFR receptor in commercial potato (Solanum tuberosum) genotypes susceptible to R. solanacearum. In these studies, we demonstrate that EFR expression confers resistance to bacterial wilt in the potato cultivar INIA Iporá under controlled conditions in growth chambers, with a significant reduction in the progression of the disease (Boschi et al., 2017). Furthermore, it was confirmed that these genotypes show resistance under conditions similar to natural infection in the field. We evidenced a restricted colonization of the pathogen at the vascular level compared to non-transformed controls, which may explain the greater resistance to the disease (Fort et al., 2020).
Some reports indicate that the increase in immunity can be accompanied by performance penalties varying between 4% and 21% depending on the culture and the defense characteristic expressed (Heil, 2002; Ning et al., 2017).
The objective of this work was to evaluate the productive performance of two INIA Iporá potato transformation events with the EFR gene with high resistance to R. solanacearum (3 and 12) in the absence of the pathogen. Under biosafety standards and during two cultivation cycles, agronomic and yield characteristics were analyzed. This research contributes towards the generation of potato genotypes resistant to bacterial wilt of agronomic interest through the use of the EFR gene.
Materials and Methods
Genotypes evaluated and culture conditions
The commercial potato cultivar INIA Iporá, a high-yielding cultivar that does not possess genes for resistance to bacterial wilt, -like all commercially available potato cultivars-was used. It is characterized by producing a high number of stems per plant (5 to 8), with high productivity compared to other potato cultivars (ENC, INASE, 2011-2020) (ENC Instituto Nacional de Semilas, 2021). This genotype was transformed with the EFR receptor gene as previously described by Boschi et al. (2017). Two events that showed high resistance to R. solanacearum, INIA Iporá EFR 3 and INIA Iporá EFR 12 (Boschi et al., 2017), were selected for this evaluation.
The plants were micropropagated in vitro from nodes maintained in Murashige and Skoog medium with sucrose 30 g/l and maintained at 22°C with cycles of 16:8 h of light:dark. Two weeks later, the seedlings were transferred to nurseries containing commercial peat (TREF mix) and kept under greenhouse conditions with natural light until the minitubers were harvested. These minitubers were then used as seed under field conditions in a field destined for this purpose for the production of seed tubers to be used in agronomic evaluation trials.
Experimental design
The study was carried out at INIA Las Brujas experimental station (latitude 34″40′ S, longitude 56″20′ W, altitude 32 m.a.s.l.), under the current regulations for the evaluation of genetically modified material according to the commission for the risk management of the Ministry of Livestock, Agriculture and Fisheries (Uruguay). The test conditions were the same (both soil and environmental) for the three genotypes analyzed: INIA Iporá untransformedwt, INIA Iporá EFR 3, and INIA Iporá EFR 12 events.
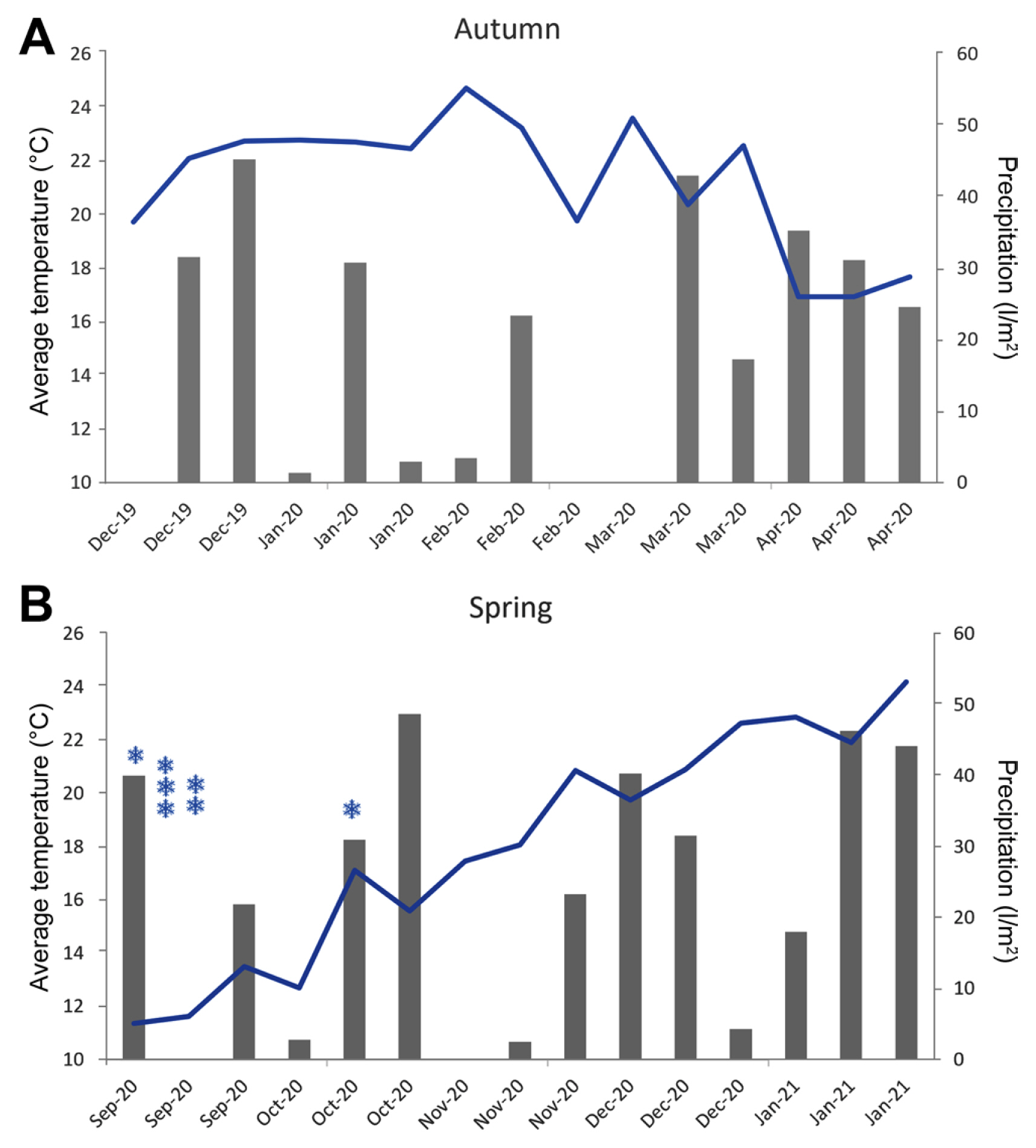
Two field trials were carried out in the autumn 2020 and spring 2020/21 harvests. The autumn trial 2020 lasted 140 days (sowing 12/4/2019, harvest 4/22/2020). The spring trial 2020/21 lasted 148 days (sowing 9/1/2020, harvest 01/27/2021). The agrometeorological conditions of these two periods are summarized in Fig. 1. The agronomic management of the trials was carried out according to the production practices for each of the seasons. A basic fertilization was applied at the time of planting with ammonium phosphate at a dose of 250 kg/ha and a refertilization with urea at a dose of 50 kg/ha. Weed management was carried out with mechanical hoes at the beginning of the crop. Later on, in specific situations, weed control was manually done. No insecticide or fungicide were applied to control pests or pathogens. Both tests were carried out with irrigation using the dripper system (one tape per bed with emitters spaced 30 cm apart and a flow rate per emitter of 1 l/h).
In both trials the experimental design was a randomized complete block with three replications. In the autumn 2020 trial, the plot had two rows spaced 0.8 m apart and 5 m long, the target plant density was 40,000 plants/ha, consisting in 5 plants per plot. All plants in the plots were analyzed. In the spring trial 2020/21 the design and conformation of the plots were the same as the autumn trial 2020, with 30 plants per plot.
Phenotype characterization
In the study of the phenotypic characterization, the guidelines of the Union for the Protection of New Varieties of Plants (UPOV), the document prepared for potato cultivation TG/23/6 (International Union for The Protection of New Varieties of Plants, 2004), were followed. The characteristics studied were: percentage of emergence measured at 30 days of cultivation, vigor evaluated at 57 and 70 days in the spring harvest and at 47 and 54 days in autumn, plant height measured in spring at 75 days and in autumn 54 days after planting. In the autumn cycle, at 54 days the observations of leaf morphology, number of stems and plant architecture were made. The number of days to flower initiation was quantified, defined when 50% of the plants had the first open flower. The vigor of the plants was described according to a scale of 1 to 9, being 1 very little vigorous and 9 very vigorous.
Performance analysis
In the yield analysis, the procedure of the National Evaluation of Potato Cultivars was followed, a mandatory requirement for the National Registry of Cultivars (ENC Instituto Nacional de Semilas, 2021). To measure total yield, all the tubers in the plot were harvested, counted, and weighted. The time of harvest was defined according to the time of senescence. The harvested tubers were classified according to tuber weight considering the following categories: commercial (greater than 80 g), seed (between 40 and 80 g), and discard (less than 40 g).
Results
Phenotype characterization. The phenotypic characterization carried out included stem length, leaf length, leaf width, leaf opening, number of stems per plant and height of the three genotypes studied in the two trials. Phenotypic traits were only evaluated in autumn 2020, and plant height of the three studied genotypes in both seasons, presented in Table 1. The main differences observed between the genotypes were in the length of the second stem, resulting in INIA Iporá EFR 12 being shorter than INIA Iporá EFR 3 and Iporá wt. In days to flowering, a trend towards later flowering was observed in transformed plants, being consistently higher in EFR 12. Likewise, a certain difference was observed in the visual appreciation of the leaf opening in EFR 12 (Fig. 2). For the other phenotypic traits, no relevant clear trends were observed.
Performance characterization. Yield analysis was performed with the two trials together (autumn and spring environment) to verify the existence of genotype-by-environment interaction. The ANOVA showed that there was significance in the model, between the genotypes, between the environments and also in the genotype (G) by environment (E) interaction. The GxE interaction was significant, which would indicate that in different environments the agronomic outcome may vary. Therefore, the analysis of each genotype in the two evaluated periods of autumn and spring was carried out separately (Table 2). In each productive cycle (autumn and spring) the total yield per area and the main components of yield, average number of tubers per plant and average weight of the tubers production were quantified for each genotype (Table 3, Fig. 2). All genotypes reached a higher total yield in the autumn trial (average yield, 34.8 t/ha) where there were favorable environmental conditions for the crop compared to the spring trial (average yield, 18.8 t/ha). In both cycles, the Iporá EFR 3 genotype exhibited significantly lower performance than the Iporá wt. Iporá EFR 12, on the other hand presented a lower yield than Iporá wt in autumn, but in spring it surpassed the untransformed control (Table 3, Fig. 3).
The difference between Iporá EFR 3, and Iporá wt in yield could be explained as a result of a lower number of tubers per plant produced in the transformed genotype in both cycles. No significant differences were observed in the mean weight of the tubers between Iporá EFR 3 and wt. However, the difference in yield between Iporá EFR 12 and Iporá wt in the autumn cycle was explained by the lower average weight of the tubers compared to the Iporá wt control. In the spring cycle, on the other hand, Iporá EFR 12 did not present significant differences in yield or its components with respect to the untransformed control. Complementing the analysis, Table 4 shows the distribution of the number of tubers per plant and its representation by category in each trial. The average number of tubers per plant in Iporá EFR 3 was lower, which explains the reduction in yield in both trials. This effect was not observed for Iporá EFR 12, which has a slightly higher number of tubers than Iporá without transformation in the spring trial. These data seem to infer that while tuber differentiation was affected in Iporá EFR 3, in Iporá EFR 12 the filling of the tuber was mainly affected in the autumn trial, resulting in the highest percentage of tubers in the lowest category observed for this genotype. while in the spring trial, EFR 12 did not show difficulty in filling, obtaining a result even higher than the wt.
During the autumn cycle, the incidence of skin disease in potato tuber, known as ‘scabies’ caused by Streptomyces scabies in transformed events, was observed. In Iporá EFR 3 it affected up to 25% of the tubers and in Iporá EFR 12 it was less than 5%, while it was not observed in the non-transformed one. The incidence of this disease is normally higher in the autumn cycle due to cool temperatures and rainfall (Fig. 1). During the spring cycle, the disease did not appear.
Discussion
Several authors reported in different plant species the positive effect of the use of the EFR gene on the control of bacterial pathogens in controlled conditions (Kunwar et al., 2018; Lacombe et al., 2010; Mitre et al., 2021; Pfeilmeier et al., 2019; Schoonbeek et al., 2015; Schwessinger et al., 2015). Previously, our research group reported the effect of the EFR gene in controlling the development of bacterial wilt in potato (Boschi et al., 2017; Fort et al., 2020). In this work, we evaluated the effect of the efr gene on agronomic aspects and performance in the absence of disease, advancing in the search for resistant germplasm of commercial interest.
Regarding phenotypic characteristics, differences were only observed in the transformed events with respect to the untransformed variety in the leaf opening and in the length of the second stem (Table 1). Leaf thickness and habit are very important elements in defining performance. The more open leaves, the higher the light extinction coefficient. The dry matter production of any crop is determined by the capture and utilization efficiency of solar energy. According to Boote and Loomis (1991), the capture depends on the incident radiation, the leaf area and the extinction coefficient of light (k, directly depends on the arrangement of the leaves, insertion angle).
The productive performance of both production cycles, autumn and spring, were different, being higher in autumn (Table 3). Although in the conditions of Uruguay there is a tendency to the autumn cycle being more productive, the differences are not so pronounced (on average 5% higher according to the Farmer Observatory [Observatorio granjero]) (Mercado Modelo, 2017). In the trial, these differences were on average greater than 50% and this could be due to the high temperatures of the spring cycle that affected during the tuberization period. But, in addition, the tuberization period considered from the beginning of flowering time to harvest in autumn was 106 days, and in spring 85 days. These differences are related to the different photoperiod, temperature, heliophany and rainfall conditions that occurred during the two growing cycles, where the cooler night temperatures in autumn favored tuberization (Fig. 1). During the harvest period for both trials there was abundant rainfall that made harvesting more complex.
Yield components in potato crops comprise the average number of tubers produced and the average size of the tubers, being strongly affected by the interaction of radiation intercepted by canopy and temperature (Struik, 2007). A relevant aspect is the delay in flowering time observed in the two transformed genotypes. The peak of stolon production and the beginning of tuberization occurs during the flowering period; therefore, flowering is considered an indicator of tuber formation. The crops start the tuberization at the moment of beginning of flowering (first flower open) or just before that moment (Celis-Gamboa et al., 2003). The observed delay in flowering (5 days in EFR 3 in both cycles and 7 and 9 days in EFR 12 in autumn and spring, respectively) implies a delay in the onset of tuber filling. The longest delay of EFR 12, could explain the greater differentiation of tubers, that could cause greater competition between them, affecting the final size. According to Struik (2007), the relative variability of the size of the tubers (the coefficient of variation of the size of the tuber) is rather constant once the tuberization is complete and depends on the number of tubers, although the relation can be complex.
Iporá EFR 12 showed a high productive potential associated with a good average number of differentiated tubers per plant in both cycles (Table 4). However, the productive result obtained by Iporá EFR 12 was very different between the cultivation cycles. In autumn, a lower development of the aerial part was observed in this genotype (see plant height, vigor and height of the second stem) (Table 1), reducing the source. In addition, the delay in flowering and filling of the tubers together with greater competition between a high number of differentiated tubers could negatively affect the final size and yield. This did not happen in spring, where the development of the plants was higher, the filling of the tubers occurred in a better way, achieving a good average size of tubers and yield compared to the rest of the genotypes. Iporá EFR 3 showed a lower yield in both production cycles, mainly explained by a lower number of differentiated tubers per plant. However, the lower number reduced the competition between them and allowed a better filling of the differentiated tubers.
Kunwar et al. (2018) reported the performance of EFR transformed tomato plants in the field in the presence of R solanaceaurm. In this work, there was no performance penalty in controlled field conditions in presence of the pathogen, the yields reported reached up to a 170% higher in the transformed genotypes when compared with non-transformed plants. Analyzing the performance of each genotype, Iporá EFR 3 presented a decrease in yield close to 40% in reference to Iporá wt in both production cycles. Instead, INIA Iporá 12, while showing a reduction ~40% in autumn when compared with non-transformed genotypes, showed a yield of 150% vs. Iporá wt in the spring assay. Considering the two crop cycles, the average yield of EFR 12 maintains a higher average yield than the other genotypes evaluated. This is a preliminary observation that will have to be corroborated in successive trials. The potential of EFR-12 genotype as a resistant genotype can be highlighted, it performed better in a growth-limiting environment, under stress conditions. High temperatures in tropical climates favor the spread of the pathogen, and certainly is prevalent in tropical regions. Another favorable point is the longer vegetative period to flowering onset, as this represents higher leaf area index (LAI). The fraction of light passing through the canopy without interception by the leaves depends on the LAI which in turn determines the proportion of ground covered by green leaves. Intercepted radiation has been found to be linearly correlated with the quantity of dry matter produced which, in the case of potatoes, is distributed over the tubers and the other parts of the plant (Haverkort, 2007). Besides, EFR-12 has also longer tuberization period under environmental favorable conditions. Taking these factors into consideration (better performance under stress conditions, longer vegetative period until the onset of flowering and thus longer tuberization period) could explain a regular performance in different environments.
The performance differences observed between the two transformation events could be partially explained by the transformation methodology used (Boschi et al., 2017). Agrobacterium tumefaciens-mediated genetic transformation does not control gene copy number, and insertion site is random. It is well documented that this transformation methodology could influence the level of gene expression, as well as performance (Dong and Ronald, 2021; Kamthan et al., 2016). Precision enhancement, where the selective insertion of transgenes occurs in predetermined places in the plant genome, called genomic safe ports, is a desirable alternative with a relatively simple methodology. None of the previous evaluations of the EFR gene in different plant genetic backgrounds were carried out using precision breeding techniques that can control the copy number and the safe genomic insertion (Dong and Ronald, 2019; Dong et al., 2020), being an interesting strategy for advance in this field
Considering the contrasting yield behavior of EFR 12 during the two cycles, loss of yield was not observed and this rule out a negative impact of the EFR gene on development and productivity. According to Mitre et al. (2021) the EFR gene was evaluated in different genetic backgrounds, including proof of concept works such as commercial varieties of tomato, potato, rice, wheat, sweet orange, and resulting in a contribution in terms of control of bacterial wilt. This indicates that the EFR gene represents a contribution to resistance regardless of the genetic environment of the construct. The performance results found in Iporá-EFR highlight the importance of using transformation methods that allow precise insertion in genomic safe ports, which avoid expression in the host plant beyond that expected. The value of Iporá EFR 12 as a parental genotype is highlighted as it presents resistance in a commercial variety with a tetraploid genetic background. Additionally, the INIA Iporá cultivar presents immunity to PVY (Dalla Rizza et al., 2007), presents pollen with a low percentage of viability, and in Uruguay it is one of the cultivars most recognized by farmers for its high adaptability and production potential. The results shown both in resistance to R. solanacearum (Boschi et al., 2017), and in productivity in the two seasons make INIA Iporá EFR 12 a very interesting alternative for production sites where the presence of R. solanacearum affects crops severely or makes potato production unviable due to its serious impact. The results presented here encourage further field studies to assess the yield production of Iporá EFR genotypes in the presence of the pathogen. Even INIA Iporá 3, that showed lower production percentages in both trials could comprise an important germoplasm in endemic regions were no resistance variants are available.
Conclusion
The Iporá EFR 12 genotype in the absence of R. solanacearum presented an annual productive yield similar to INIA Iporá wt. Iporá EFR 12 is expected to maintain productive stability when bacterial wilt is present in the planting plots. This genotype could be an interesting variety for production where bacterial wilt disease is endemically present. Likewise, Iporá EFR 12 can be an alternative for the genetic base of future cultivars.
Currently, the EFR potato with bacterial wilt resistance highlights the importance of a regulatory system that can perform environmental risk assessment in countries where the disease is endemic and the potato is a staple food. The breeders of this germplasm share this advance for its use in regions that suffer from this cultural problem with prior agreement of use.
It is emphasized the convenience of making precision improvements with the EFR receptor that avoid other expressions in the genome of the host plant beyond that expected, thus facilitating regulatory aspects.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






