



The family Tylenchidae ├¢rley, 1880 is constituted by genera whose members are mostly cosmopolitan (Siddiqi, 2000). Members of this group are normally represented in any given soil sample, and are more dominant in soils rich in algae and mosses (Ferris and Bongers, 2006; Siddiqi, 2000; Yeates and Bird, 1994). Currently, the family comprises 414 nominal species belonging to 44 genera (Mortazavi et al., 2021; Panahandeh et al., 2019; Qing and Bert, 2019). This group is characterized by several ancestral characters such as weak feeding apparatus, undifferentiated non-muscular corpus and the filiform tails (Siddiqi, 2000). They do not represent economically important plant pathogens and are therefore grouped as weak plant parasites (Qing and Bert, 2017; Siddiqi, 2000). In fact, their feeding behavior is still debatable, considering the fact that food preferences of most of the described species are unknown (Munawar et al., 2021). Some members are regarded as root hair feeders while several members of Filenchus Andr├Īssy, 1954 can feed and reproduce on fungi (Bert et al., 2010; Okada et al., 2005; Siddiqi, 2000). Notwithstanding the mystery of their feeding behavior, members of this group play important ecological roles as soil health indicators (Bongers and Bongers, 1998; Munawar et al., 2021).
Filenchus is a member genus of the subfamily Tylenchinae ├¢rley, 1880 and represents one of the most diverse genera of the family Tylenchidae (Bert et al., 2010; Geraert, 2008). There is limited consensus on the actual number of nominal species that are recognized within this genus. In the review of Raski and Geraert (1987), member species of the genera Ottolenchus Husain and Khan, 1965; Dactylotylenchus Wu, 1968; Lambertia Brzeski, 1977; Duosulcius Siddiqi, 1979; Zanenchus Siddiqi, 1979 and Discotylenchus Siddiqi, 1980 were considered to be representatives of congeneric species. These genera were therefore considered to be junior synonyms of Filenchus. However, this synonymization was rejected by Siddiqi (2000). Brzeski (1998) and Andr├Īssy (2007) accepted the synonymization of Ottolenchus Husain and Khan, 1967 with Filenchus. In the recent comprehensive species compilation, Geraert (2008) maintained the synonym position of the aboved listed genera except Discotylenchus Siddiqi, 1980. Recent progresses in molecular evidence portray Filenchus as a clearly polyphyletic genus, with the four-incisured Filenchus group constituting a well-supported clade independent of the two-incisured Filenchus species (Ottolenchus) which form several separate clades (Qing and Bert, 2019; Qing et al., 2017). Therefore, the number of nominal species that are recognized within the genus Filenchus differs depending on the classification system followed. For example, Brzeski (1998) listed 79 nominal species, Siddiqi (2000) 55, Andr├Īssy (2007) 75 while Geraert (2008) comprehensively compiled a list of 94 nominal species. In addition, many of the listed member species such as F. cylindricus (Thorne and Malek, 1968) Niblack and Bernard, 1985, F. filipjevi Andr├Īssy, 1988, F. thornei (Andr├Īssy, 1954) Andr├Īssy, 1963, F. cylindricaudus (Wu, 1969), Siddiqi, 1986, F. japonicus Mizukubo and Minagawa, 1986, and F. butteus (Thorne and Malek, 1968) Raski and Geraert, 1987, among others have been subjects of synonyms by various taxonomists (Andr├Īssy, 2007; Brzeski, 1997; Raski and Geraert, 1987).
Accordingly, despite the speciose nature of the subfamily Tylenchinae, individual species delimitation remains uncertain and there is limited unanimity regarding the taxonomic position of various species and genera (Andr├Īssy, 2007; Bai et al., 2020; Brzeski, 1997; Siddiqi, 2000). As noted by De Ley and Blaxter (2002), it is clear that microscopical data alone cannot provide adquate diagnostic resolution required to consistently delineate species that possess limited morphological differentiating characters. Integrative identification considering both morphological characters and molecular phylogenetic inferences provides a better supported approach in species designation even though delineation based on DNA is still, depending on taxonomic opinions, a controversial matter (see Fortuner et al., 2018; Mwamula et al., 2020a, 2020b, 2020c, 2021; Subbotin et al., 2005). Unfortunatley, DNA sequence data of many of these species is still unavailable. Filenchus cylindricus has been consistently described from rhizosphere of various plants including grasses (Poaceae and Gramineae like maize), potato, onion, garlic, common guava and citrus, among others (Dobrin and Geraert, 1994; Elmiligy, 1971; Thorne and Malek, 1968; Zeidan and Geraert, 1991), and is therefore, a plant-parasitic nematode albeit with non-substantive pathogenicity level. Following our taxonomic studies on turfgrass parasitic nematodes in Korea, a population of F. cylindricus was found associated with irregular discoloration and patchy thinning of Poa pratensis on putting greens of Gapyeong Benest golf club. Herein, we provide molecular characterization of the species, and offer additional update of morphometric and morphological data. Also, the current characterization represents the first record of the species in Korea.
Materials and Methods
Nematode population and extraction
Nematode population was extracted from sod samples taken from Gapyeong Benest golf club. The sample was taken from putting greens, composed of mainly loamy sand soils, with Poa pratensis as the host crop. Nematodes were extracted from the sod cores using a combination of modified CobbŌĆÖs sieving and Baermann funnel method (Jenkins, 1964). Nematode specimens belonging to the genus Filenchus were handpicked from the nematode suspension under a Nikon SMZ 1000 stereomicroscope (Nikon, Tokyo, Japan); and were subsequently characterized morphologically and molecularly.
Morphological characterization
Thirty-six female and 22 male specimens from sample KP4 were killed, fixed, and processed to pure glycerin (Seinhorst, 1959). Photomicrographs and morphometric data were taken using a Zeiss imager Z2 microscope (Carl Zeiss, Jena, Germany) fitted with Axio-vision software: Material Science Software for Research and Engineering software (Carl Zeiss). Species identification was made following the identification key and species descriptions compiled by Brzeski (1997), Geraert (2008), and Raski and Geraert (1987).
Molecular characterization
Ribosomal DNA was extracted from single female specimens using the DNA extraction kit WizPure, as described by Iwahori et al. (2000). The primer set D2A (5ŌĆ▓-ACA AGT ACC GTG AGG GAA AGT TG-3ŌĆ▓) and D3B (5ŌĆ▓-TCG GAA GGA ACC AGC TAC TA-3ŌĆ▓) (Nunn, 1992) was used to amplify the D2-D3 expansion segment of 28S-rRNA while TW81 (5ŌĆ▓-GTT TCC GTA GGT GAA CCT GC-3ŌĆ▓) and AB28 (5ŌĆ▓-ATA TGC TTA AGT TCA GCG GGT-3ŌĆ▓) (Curran et al., 1994) amplified the ITS (internal transcribed spacer)-rRNA gene. Polymerase chain reaction (PCR) was performed with a PCR cycler (T100, Bio-Rad, Hercules, CA, USA), and the program was set as follows: initial denaturation at 95┬░C for 5 min, 35 cycles at 95┬░C for 30 s, annealing at 53┬░C for 30 s, 72┬░C for 1 min 10 s, and one cycle at 72┬░C for 5 min. The PCR products were purified using a PCR purification kit (Qiagen, Hilden, Germany), and quantification was done using a quickdrop spectrophotometer (Molecular Devices, San Jose, CA, USA). The purified products were subsequently directly sequenced in both directions using the same primers. DNA sequencing was conducted by Macrogen. The obtained DNA sequences were submitted to the GenBank database under the accession numbers MN752397 and MN752398 (28S-rRNA), and MN752395 and MN752396 (ITS-rRNA).
Phylogenetic analysis
The obtained sequences of 28S-rRNA and ITS-rRNA were aligned using ClustalX 1.83 (Thompson et al., 1997) along with the corresponding comparable sequence datasets of genetically close species within the genus Filenchus and other related genera published in GenBank (Atighi et al., 2013; Mortazavi et al., 2021; Munawar et al., 2021; Panahandeh et al., 2019). Neothada cancellata and Basiria bhabi were used as the outgroup taxa for 28S-rRNA and ITS-rRNA sequence datasets, respectively. The generated alignments were then analyzed with Bayesian inference using MrBayes 3.2.6 (Ronquist et al., 2012). The best-fit model of nucleotide substitution was achieved using jModelTest v2.1.7 (Darriba et al., 2012). The general time reversible substitution model with estimation of invariant sites and assuming a gamma distribution with four categories (GTR + I + G) was taken as the appropriate nucleotide substitution model for the two analyses. Bayesian inference analysis for each gene was initiated with a random starting tree, and run with four chains for 1 ├Ś 106 generations. Posterior probabilities were estimated using the Markov chain Monte Carlo method, and consensus trees were generated with 50% majority rule. Trees were subsequently visualized and edited using FigTree software v1.4.4. Intraspecific and interspecific genetic variations were checked by PAUP* v4.0a169 (Swofford, 2003).
Results
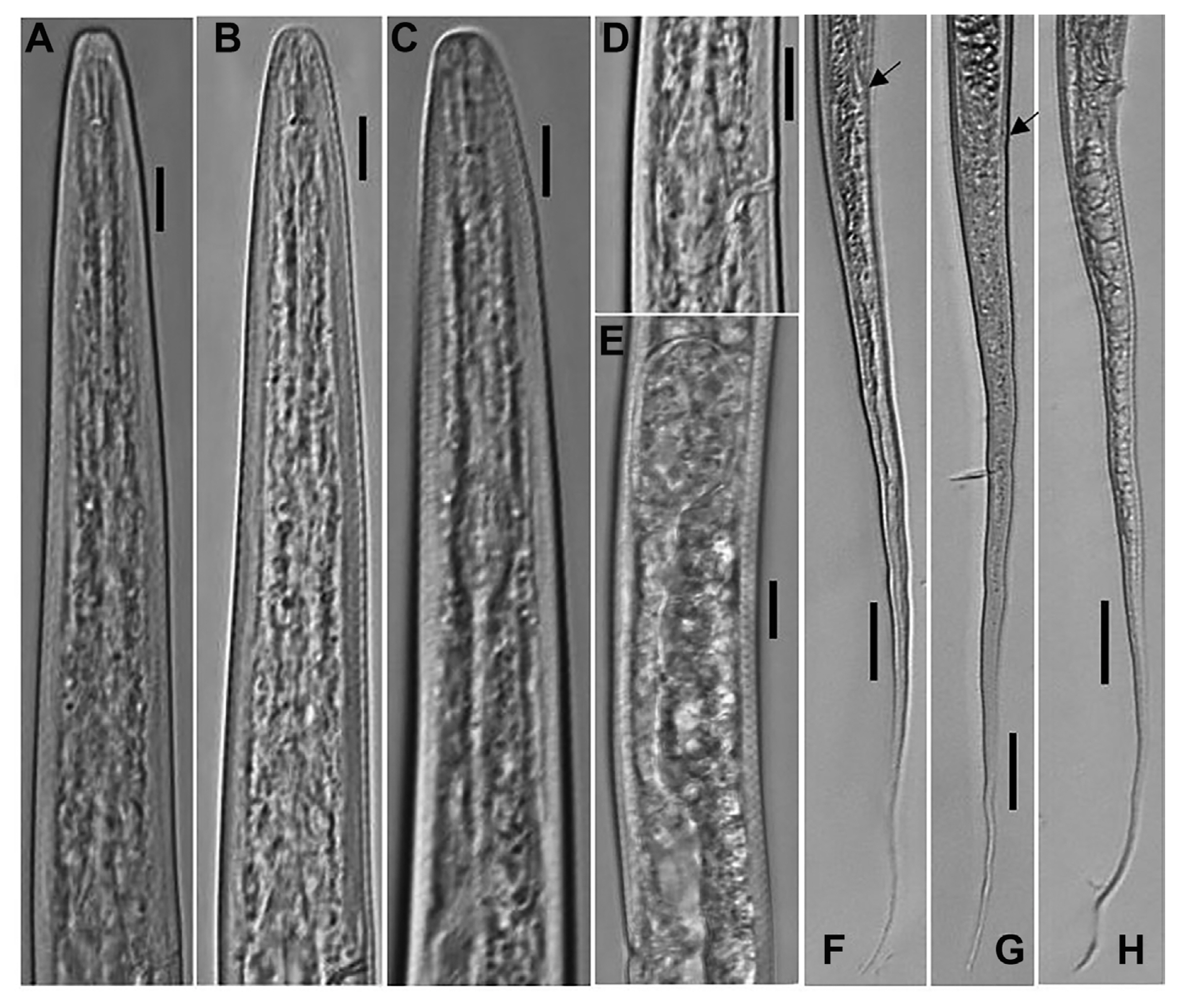
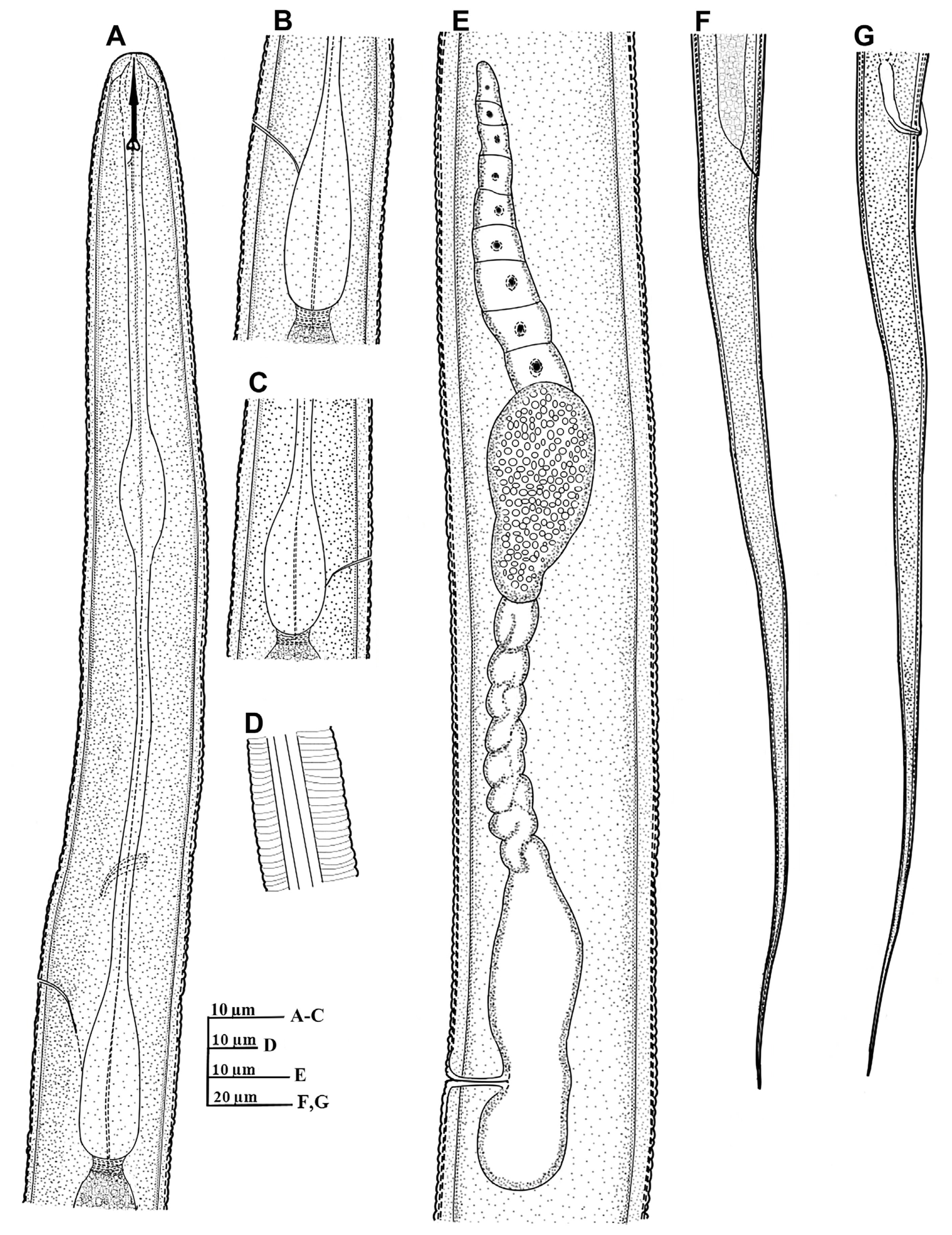
Filenchus cylindricus (Thorne and Malek, 1968) Niblack and Bernard, 1985 (Figs. 1 and 2)
Description
Female
Body about 1 mm, straight to slightly ventrally-curved when heat killed. Maximum body width 24.0-27.0 ╬╝m; vulval body width 21.0-26.0 ╬╝m. Cuticle comprised of fine annuli, about 1.5 ╬╝m wide at mid-body. Head annuli obscure under light microscope, visible in some specimens. Lip region 3.3 (3.0-4.0) ╬╝m high, round to anteriorly flattened, continuous with the body contour, and 6.9 (6.5-7.5) ╬╝m wide. Stylet well developed, stylet knobs distinct and round, often sloping posteriorly. Dorsal gland orifice located at 1.6-2.7 ╬╝m posterior the stylet knob base. Esophagus with elongated, oval median bulb, 6.3 (5.0-7.0) ╬╝m wide, with small, faintly cuticularized valve, located at 29.0-39% of the esophagus length. Nerve ring encircling a portion of posterior half of isthmus. Secretory-excretory pore located approximately mid to the upper part of the basal bulb region. Hemizonid ca 1-2 annuli anterior to excretory pore. Basal bulb elongated and pyriform, 8.6 (7.0-11.0) ╬╝m wide and 24.4 (22.0-33.5) ╬╝m long. Lateral field with four incisures, covering about 1/4-1/5 body width. Ovary single, outstretched, spermatheca offset, elongated, 28.2 (19.0-37.5) ╬╝m long, filled with round sperm. Vagina occupying 37-55% of corresponding body width. Post vulval uterine sac prominent, 14.0 (13.0-16.0) ╬╝m long; occupying 50-71% of vulval body width. Vulva to anus length long, 0.8-1.2 times the tail length. Anal body width 15.0-17.5 ╬╝m. Tail long, tapering gradually in the anterior two thirds, becoming filiform in the distal portion, terminating in a finely rounded or pointed terminus. Tail annuli indistinct in the posterior filiform part.
Male
Habitus generally similar to female but slender (Table 1). Body generally straight when heat killed and after fixation. Cuticle with fine annuli, about 1.0-1.5 ╬╝m wide at mid-body. Stylet well developed, with distinct round stylet knobs. Testis outstretched. Spicules arcuate, gubernaculum trough-shaped, slightly curved. Bursa weakly developed, with crenate margins, extending from base of spicules to about one spicule length past the cloaca. Tail long, tapering gradually and filiform in the distal part.
Remarks
The current studied population is identical to the Colorado population (Raski and Geraert, 1987), F. cylindricus syn. T. hageneri population (Elmiligy, 1971) and the paratypes (Brzeski, 1997), both in morphological characters and morphometrics (Table 1), but slightly differs from the Sudan population (Zeidan and Geraert, 1991) by a relatively longer body length (0.94-1.1 vs. 0.70-0.87 mm), longer esophagus (138.0-152.0 vs. 116-127 ╬╝m long), longer distance from anterior end to secretory-excretory pore (115-129 vs. 87-103 ╬╝m), longer vulva to anus distance (184.5-267.0 vs. 112-166 ╬╝m), longer tail length (185.5-247.0 vs. 140-178 ╬╝m), and T/VA (0.8-1.2 vs. 1.0-1.4). The current population is also comparably similar to a population from Pakistan (Erum and Shahina, 2010), except for a higher a ratio (37.2-44.7 vs. 28-32). However, the morphometrics of the population from South Carolina (Zeng et al., 2012) and the recently characterized population from Southern Alberta, Canada (Munawar et al., 2022) significantly differ from the current studied population and other characterized populations including the holotype and paratypes; mainly by shorter body length (531.0-560.5 and 557.0-711.0 ╬╝m), shorter vulva to anus distance (77.5-86.0 and 98.0-139.0 ╬╝m), shorter tail (95.8-116.2 and 112.0-159.0 ╬╝m), shorter distance from anterior end to excretory pore (80.0-82.0 and 75.0-86.0 ╬╝m), shorter pharynx (92.0-98.5 and 91.0-103.0 ╬╝m), respectively, and lower a ratio (23.1-26.0) in Zeng et al. (2012). The current population characterization represents the first record of F. cylindricus in Korea and therefore extends its distribution to seven countries (USA: Thorne and Malek, 1968; Sudan: Zeidan and Geraert, 1991; Bulgaria: Katalan-Gateva and Tso, 1977; Romania: Dobrin and Rosca, 1996; Turkey: Erdogus et al., 2010; Pakistan: Erum and Shahina, 2010; Canada: Munawar et al., 2022; Korea).
Molecular characterization and phylogenetic relationships
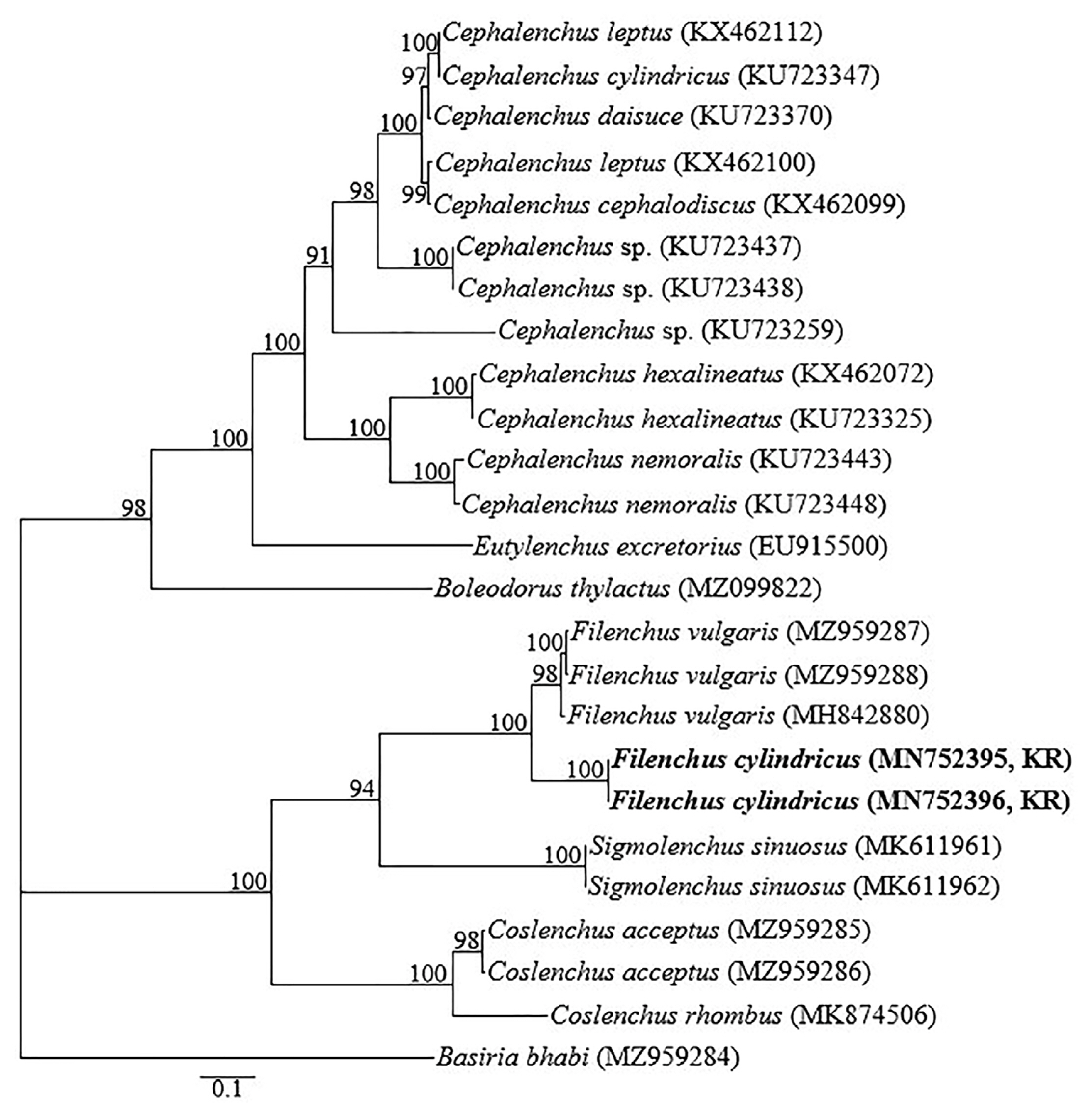
The D2-D3 expansion segment of 28S-rRNA, and ITS-rRNA gene yielded single fragments of approximately 700 and 800 bp, respectively based on gel electrophoresis estimations (data not shown). The top five D2-D3 expansion segment BLASTN hit for the two sequences (MN752397 and MN752398) obtained from a population confirmed morphologically as F. cylindricus included F. sindhicus Shahina and Maqbool, 1994 isolate 11T05J09 (JQ005012), F. thornei (Andr├Īssy, 1954) Andr├Īssy, 1963 isolate 21T16I09 (JQ005014), F. annulatus (Siddiqui and Khan, 1983) Siddiqi, 1986 isolate 3T08B11 (JQ005017), F. quartus (Szczygiel, 1969) Lownsbery and Lownsbery, 1985 isolate 1T28I09 (JQ005016) and unidentified species, Filenchus sp. isolate 6T11G09 (JQ005015), differing by 9-10 bp (1.2-1.3%), 14-15 bp (1.9-2.0%), 37-38 bp (5.0-5.1%), 38-39 bp (5.1-5.3%), and 40-41 bp (5.4-5.6%), respectively. Intraspecific variation within the two generated F. cylindricus sequences was only 1 bp. The two ITS sequences of F. cylindricus (MN752395 and MN752396) showed a relative level of homology to F. vulgaris (MH842880, MZ959287, and MZ959288), the only ITS sequences of Filenchus available in GenBank, from which differences of 140-154 bp (18.9-20.1%) were recorded. Forty-two 28S-rRNA and 25 ITS-rRNA sequences of various species within the genus Filenchus and other related genera constituted the datasets for phylogenetic analysis. Phylogenetic relationships, as inferred from Bayesian analysis of the dataset with GTR + I + G substitution model are shown in Figs. 3 and 4.
Discussion
In the 28S-rRNA analysis, the 50% majority-rule consensus tree from Bayesian analysis generated with the GTR + I + G model agrees with the analysis by Munawar et al. (2021). Filenchus cylindricus and all other Filenchus sequences except F. discrepans (KX156317) formed a highly supported clade (posterior probability [PP] = 100%) albeit with some Tylenchus species. This is not unanticipated since the two genera are known to be more similar morphologically and molecularly than with other related genera (Bert et al., 2008; Geraert, 2008; Holterman et al., 2008; Raski and Geraert 1987). Notably, F. discrepans (KX156317) occupied an independent position of the tree. This agrees with Mortazavi et al. (2021) regarding the polyphyletic nature of the members of the subfamily Tylenchinae. In ITS-rRNA analysis, the two F. cylindricus sequences were grouped together with the only available ITS-rRNA sequences of Filenchus (F. vulgaris) in GenBank at the time of the current study.
Specifically, the grouping of F. cylindricus in a highly supported subclade (PP = 100%) with F. sindhicus and F. thornei is of interest, and reaffirms the significance of integrative identification of morphologically close species considering both morphological characters and molecular phylogenetic analysis. Filenchus cylindricus was first described as Tylenchus cylindricus from prairie sod in South Dakota, and Holbrook, Nebraska by Thorne and Malek (1968). Raski and Geraert (1987) then reviewed the genus and synonymized T. hageneri Elmiligy, 1971 with T. cylindricus before transferring the species to the genus Filenchus. In the same review, Raski and Geraert (1987) demonstrated that the micrographs of F. cylindricus showed no difference from F. thornei. Brzeski (1997) then synonymized F. cylindricus and other related species (F. cylindricaudus Wu. 1969, F. japonicus Mizukubo & Minagawa, 1986 and F. filipjevi Andr├Īssy, 1988) with F. thornei (Andr├Īssy, 1954) Andr├Īssy, 1963. These synonymizations were not recognized by Siddiqi (2000) and Geraert (2008), except for F. japonicus in the compilation done by Geraert (2008). In the list of Andr├Īssy (2007), F. cylindricus is listed as a synonym of F. butteus (Thorne and Malek, 1968) Raski and Geraert, 1987. In the current analysis, F. cylindricus D2-D3 expansion segment is distinct from that of F. thornei by 14-15 bp, thus confirming the independent taxonomic position of the two species. The current F. cylindricus population and other studied isolates especially the Sudan population are indeed close to F. thornei. However, F. cylindricus can be distinguished from F. thornei by its longer stylet (12-13 vs. 9-12 ╬╝m), and the characteristic longer distance from vulva to anus, translating into a lower T/VA ratio (0.8-1.4 vs. 1.4-1.8). And from F. sindhicus (the closest species based on 28S-rRNA (98% identity) by its longer stylet (12-13 vs. 7-8 ╬╝m), lower T/VA ratio (0.8-1.4 vs. 2.1) and non-hooked tail terminus vs. hooked.
The recent molecular characterization of F. cylindricus population from Southern Alberta, Canada (Munawar et al., 2022) denotes a possible existence of a species complex in form of cryptic species. Based on the 28S rRNA gene, F. cylindricus was shown to cluster with F. hazenensis in a separate clade. A review of all the population descriptions of F. cylindricus shows a wide variation in the morphometric data. For example, body length varies from 531 to 1150 ╬╝m (shortest individual in the South Carolina population [Zeng et al., 2012] and longest in the Colorado populations [Raski and Geraert, 1987]), vulva to anus distance of 77.5-267.0 ╬╝m (shortest in the South Carolina population (Zeng et al., 2012) and longest in the current population characterization), tail length of 96.0-247.0 ╬╝m (lowest value recorded in South Carolina population [Zeng et al., 2012] and highest in the current characterization and the Colorado population [Raski and Geraert, 1987]), and pharynx length of 91.0-175.0 ╬╝m, being shortest in the Southern Alberta population (Munawar et al., 2022) and longest in the Colorado population (Raski and Geraert, 1987). There is no doubt that species of this genus display intraspecific morphological variations between and within populations. However, the wide ranges recorded, especially between independent populations make species differentiation from closely related species difficult. It is thus probable that some of the wide morphometric variations within F. cylindricus, represent distinct species as evidenced by the current molecular evidence. Doblin and Geraert (1994) considered the two small-bodied populations (600-710 ╬╝m) from Romania as F. cylindricus based on the similarity of stylet length but noted that contrary to the previously reported F. cylindricus populations, individuals of these populations were significantly smaller. Moreover, their scanning electron microscope micrographs showed smooth head, different from that in Raski and Geraert (1987). It is therefore likely that the small bodied populations represent a cryptic species, and thorough morphological and molecular studies on topotype population are required for proper delineation of this species.
It is important to note however, that general morphological delineation of many other species within this speciose genus is undoubtedly challenging. This is due to the high morphological plasticity at species level, with overly indistinctive character overlaps within closely related species. The small body size coupled with lack of clearly homologous morphological characters complicates the identification of the member species of this group (Bongers and Bongers, 1998; Qing and Bert, 2019; Siddiqi, 2000). The species review and list by Geraert (2008), is full of assumed synonyms suggested by various taxonomists in the past years. There are generally divided opinions regarding their classification from species level up to as far as family level. Even with the current advancements in molecular techniques applied in taxonomic studies, species of this family have not been given the necessary attention since more emphasis is always attributed to the most destructive nematode species of agricultural importance. In conclusion, there is need to molecularly characterize and compare type specimens or populations from type localities with other studied populations of the species of this genus. This will increase taxonomic resolution and consensus among biologists regarding the taxonomic positions of the contested species and (or) populations.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






