



Bacterial wilt of bean, caused by Curtobacterium flaccumfaciens pv. flaccumfaciens (CFF), is an economically important disease in commercial dry edible bean production systems worldwide (Osdaghi et al., 2016), including Western Canada (Huang et al., 2009). The bacterium is a small, infectious, Gram-positive, rod-shaped prokaryote that colonizes xylem tissues impeding water movement, which causes wilting symptoms in host plants, or blank areas in the canopy when plants are killed. In addition to colonizing the xylem vessels, CFF can also infect developing seeds and become seed-borne (Huang et al., 2007). Once colonization of the vascular elements has occurred, there are no effective control measures, therefore management of the disease is accomplished preventatively by use of clean, disease-free seed, application of bactericidal seed treatments, and crop rotation. Bacterial wilt on bean has become more serious in recent years and its resurgence has raised concerns regarding how the disease will be managed in production areas around the world (Harveson et al., 2015; Huang et al., 2009; Osdaghi et al., 2016, 2020). The Great Northern market class of dry edible bean can display discoloration or staining of the white seed coat caused by the colored pigments produced by CFF infection. CFF is known to produce colored pigments, and staining that can be yellow, orange, pink, red or purple, depending on the variant (Osdaghi et al., 2016). These staining symptoms have been reported to be negatively correlated with percent seed emergence (Huang et al., 2007).
Many plant pathogenic bacteria and fungi form biofilms at one or more stages of their disease cycles (Harding et al., 2010; Padmavathi et al., 2017; Ramey et al., 2004; Velmourougane et al., 2017). Biofilms are surface-associated colonies of cells that are encased in a slimy, self-produced, polysaccharide matrix. The slimy matrix allows cells to attach to one another, and to substrates, including the cell walls of host plants, and provides a number of protective benefits that solitary cells do not enjoy (Reichhardt and Parsek, 2019). A number of biofilm-associated phenotypes have been described, the most common being increased tolerance to physical and chemical treatments that would be lethal to populations of comparable solitary or planktonic cells (Stewart et al., 2019). For example, biofilms may be up to 1,000-times more tolerant to chemical treatments than solitary or free-floating planktonic bacterial cells that lack the protection afforded within the biofilm (Gilbert and McBain, 2001). The formation of microbial biofilms as part of the etiology of plant diseases has already been demonstrated for a number of xylem-colonizing bacteria (Castiblanco and Sundin, 2016). Examples include Goss’s wilt on corn caused by Clavibacter michiganensis subsp. nebraskensis (Botti-Marino, 2017; Harding et al., 2011), bacterial wilt on many crops caused by Ralstonia solanacearum (Mori et al., 2016), ring rot on potato caused by Clavibacter michiganensis pv. sepedonicus (Howard et al., 2015), fire blight on fruit trees caused by Erwinia amylovora (Koczan et al., 2009), Stewart’s wilt of corn caused by Pantoea stewartii (Koutsoudis et al., 2006), Pierce’s disease of grape caused by Xylella fastidiosa (Marques et al., 2002), and crown gall caused by Agrobacterium tumefaciens (Tarbah and Goodman, 1987). In some instances, the ability to form biofilms is an essential virulence factor (Koczan et al., 2009; Mori et al., 2016).
Due to the important effects of biofilm formation on pathogenicity and virulence, the ability to culture and study biofilms is particularly important. However, culturing biofilms can be challenging when compared to more standard microbiological culture methods for planktonic cells, such as growth in nutrient rich broths or on semi-solid agar gels. Standard methods for culturing and evaluating biofilms are now more widely utilized (Harding and Daniels, 2017), and one method, the MBEC (minimum biocide eradication concentration) Assay method, allows rapid culturing of biofilms in a high throughput platform (Fig. 1). Using this multi-well plate-based method, one can easily determine the minimum concentration required to eradicated both biofilm and planktonic populations in the same reactor, making it especially useful for evaluation of chemical treatments and simultaneous comparison of treatment efficacy against both biofilm and planktonic cells (Harrison et al., 2010). For example, the MBEC Assay method has been used to characterize C. sepedonicus biofilms (Howard et al., 2015), C. michiganensis biofilms (Harding et al., 2011), and recently to identify novel, anti-biofilm chemicals such as oxidized silver compounds and optimal fungicide + metallic ion tank-mix partners (Harding et al., 2019). The availability of this assay was instrumental in developing the first American Society for Testing and Materials (ASTM) standard method for evaluation of disinfectant efficacy in a static reactor (ASTM Standard Method E2799-17).
The re-emergence of bacterial wilt inspired a biofilm approach to better understand the disease cycle of CFF, and to evaluate the efficacy of five bactericidal compounds. Copper, silver, and streptomycin are all known to have bactericidal effects. In this study, the efficacy of oxidized silver compounds was evaluated and compared with copper and streptomycin. The purposes of this study were to evaluate effects of CFF infection on emergence of dry bean seed, determine whether CFF colonization of dry bean tissues had a biofilm morphology and to measure the efficacy of five bactericidal chemicals (copper sulfate, copper hydroxide, oxysilver bisulfate, oxysilver nitrate and streptomycin) against CFF, and compare the sensitivity of planktonic CFF cells to those within a biofilm. It was hypothesized that CFF would form robust biofilms at some point during the disease cycle, such as vascular colonization and occlusion, and that the bacterial cells within biofilms would be recalcitrant to bactericidal inactivation when compared with planktonic cells. Furthermore, it was hypothesized that oxidized silver compounds would be effective at reducing biofilm and planktonic populations.
Materials and Methods
Organisms, culture media and emergence bioassay
Cultures of CFF were initiated from a freezer stock of an isolate obtained from a symptomatic dry bean plant field in southern Alberta in 2005 (Fig. 2). The bacterium was identified as CFF as it was a small, Gram-positive rod that formed yellow pigmented colonies on modified CNS agar and was not soluble in 3% KOH. The frozen stock was revived by streaking onto brain heart infusion (BHI) agar (BD Biosciences, San Jose, CA, USA) and incubating for 48-72 h at 22 ± 2°C at 75% relative humidity. Colonies were used to prepare inoculum on BHI agar plates or in BHI broth (BD Biosciences).
Dry bean seed (cv. AC Polaris) was obtained from fields with a history of bacterial wilt symptoms. The seed lot used in this study was colonized by a yellow variant, and the seed lot was divided into four categories based on visual appearance. To confirm the negative impact of seed infection with emergence, the symptomatic seed was sorted according to the following categories: (1) yellow discoloration and shrivelled, (2) yellow discoloration, not shrivelled, (3) brown discoloration, and (4) no discoloration. Emergence bioassays were performed in 500 ml moistened potting medium, Pro-Mix (Premier Tech Home and Garden, QC, Canada) dispensed into 15 cm diameter pots. Five seeds were sown in each pot with five biological replicates for each category. The 20 pots were arranged in a completely randomized design in a greenhouse set at 25 ± 2°C for 18 h with light and 18 ± 2°C for 6 h in the dark. Emergence counts were taken weekly, and the final emergence (after 4 weeks) was reported as the percent of seedlings emerged. This experiment was repeated two more times with similar results.
Scanning electron microscopy
Five seeds from category 2 (yellow discoloration, not shrivelled) were selected for scanning electron microscopy (SEM). Additionally, 6-week-old plants grown from category 2 seeds were evaluated by excising 1-4 cm2 samples of infected plant tissue with a scalpel or razor blade. Sample fixation was performed using a standard approach (Bozzola and Russell, 1992) as follows; sample tissues were covered with primary fixative (3% glutaraldehyde + 1.6% paraformaldehyde in 0.2 M sodium cacodylate buffer, pH = 7.5) and incubated at room temperature for 2 to 4 h. Tissues were rinsed three times (15 min each) in 0.2 M sodium cacodylate buffer. Tissues were then added to 1-3 ml (or just enough to cover plant tissues) of 1% osmium tetroxide and incubated at room temperature for at least 1 h. Rinsing was repeated and tissues were dehydrated through a graded ethanol series (serial rinses for 10 min each at the following concentrations: 35%, 50%, 70%, 85%, 95%, and 100%). Samples were post-fixed with osmium tetroxide and critical point dried. All samples were mounted on aluminium stubs, sputter coated and scanned in a Philips C-60 ESEM (Philips/FEI Co., Hillsboro, OR, USA).
Minimum biofilm eradication concentration
Culturing of biofilms and planktonic cells, and disinfectant efficacy testing, was performed in accordance to the methods and procedures outlined in ASTM E2799-17 (Standard Test Method for Testing Disinfectant Efficacy against Pseudomonas aeruginosa Biofilm) (ASTM International, 2017), using the MBEC assay device, with modifications as previously described in (Harding et al., 2011). Briefly, the bacterium was grown in BHI broth and biofilms were cultured by inoculating each well of a 96-well MBEC plate with 150 μl of CFF inoculum made from a broth culture of 105 colony forming units per ml (cfu/ml) in OSB and incubated on a humidified gyrorotary shaker at 110 rpm at 25 ± 2°C for 24 h. The biofilms were exposed to a 96-well biocide challenge plate prepared separately for each biocide. Each challenge plate included 10 replicates at each of eight concentrations of one biocide (80 wells), sterility control (3 wells), neutralizer function control (2 wells), neutralizer toxicity control (2 wells), non-treated control (4 wells), and growth control (5 wells). The MBEC assay plate lids, with biofilms attached, were placed into the biocide challenge plates for 30 min. They were then serially transfered to a sterile distilled water rinse plate (200 μl per well) for 1-2 min, a neutrlizer plate (200 μl per well) for 24 h, and then placed into sonicator for 30 min to disaggregate the biofilms. A serial dilution and spread plate on OSA were performed from each well to determine cfu/ml. For minimum biocidal concentrations (MBC) calculations, 20 μl volumes from each well of the challenge plates were transferred to a new plate containing neutralizer and enumerated as described as described above with the exception that sonication to disagregate the cells was not required. All broth, agar and media ingredients were obtained from (BD Biosciences) with the exception of the universal neutralizer solution which was prepared as follows: 1.0 g L-histidine (Sigma, St. Louis, MO, USA), 1.0 g L-cysteine (Sigma), 2.0 g reduced glutathione (Sigma) in 20 ml double distilled water filtration sterilized through 0.22 μm diameter pore size filter (Corning Inc., Kaiserslautern, Germany) and stored at −20°C. MBEC Assay plates were provided by Innovotech Inc. (Edmonton, AB, Canada). The concentrations of bactericidal chemicals were based on preliminary efficacy results (not shown). The chemicals were selected primarily to evaluate the efficacy of oxidized silver compounds against CFF biofilms with copper compounds and streptomycin as standards for comparison. Stock solutions were prepared by adding the raw product into sterile water to reach the desired concentration as follows: copper hydroxide (2,000 μg/ml), copper sulfate (2,000 μg/ml), oxysilver bisulfate (1,000 μg/ml), oxysilver nitrate (1,000 μg/ml), and streptomycin (1,024 μg/ml). Solutions were mixed thoroughly and stored at 4°C until used. Stock solutions were serially diluted by 50% by combining 0.4 ml of solution with 0.4 ml of sterile water.
Copper hydroxide, copper sulfate and streptomycin were obtained from Sigma Aldrich (Oakville, ON, Canada), oxysilver nitrate and oxysilver bisulfate were provided by Innovotech Inc.
Calculation of EC50 and EC95 and statistical analyses
The effective concentrations to cause 50% inhibition (EC50) and 95% inhibition (EC95) were calculated using Quest Graph EC50 Calculator (https://www.aatbio.com/tools/ec50-calculator) (AAT Bioquest, 2019). Student t-tests (P ≤ 0.01) were performed in Microsoft Excel 2016 (Mississauga, ON, Canada). ANOVA was performed using Statistics Kingdom online calculator (https://www.statskingdom.com/180Anova1way.html, 2022).
Results
Effects of CFF infection on dry bean emergence
Seedlings emerged from the soil 5 to 10 days after sowing. The category 1 seed (completely yellow and shrivelled) had the lowest emergence (0%), followed by category 2 (some yellowing), which had 20% emergence, while category 3 (brown discoloration) had 44% emergence, and category 4 (no disoloration or shrivelling) had the highest emergence (100%) (Fig. 3). The differences between the means of each category were statistically significant (P ≤ 0.01) according to pair-wise comparisons using a Student’s t-test (Table 1).
SEM imaging of CFF on seeds and in stems
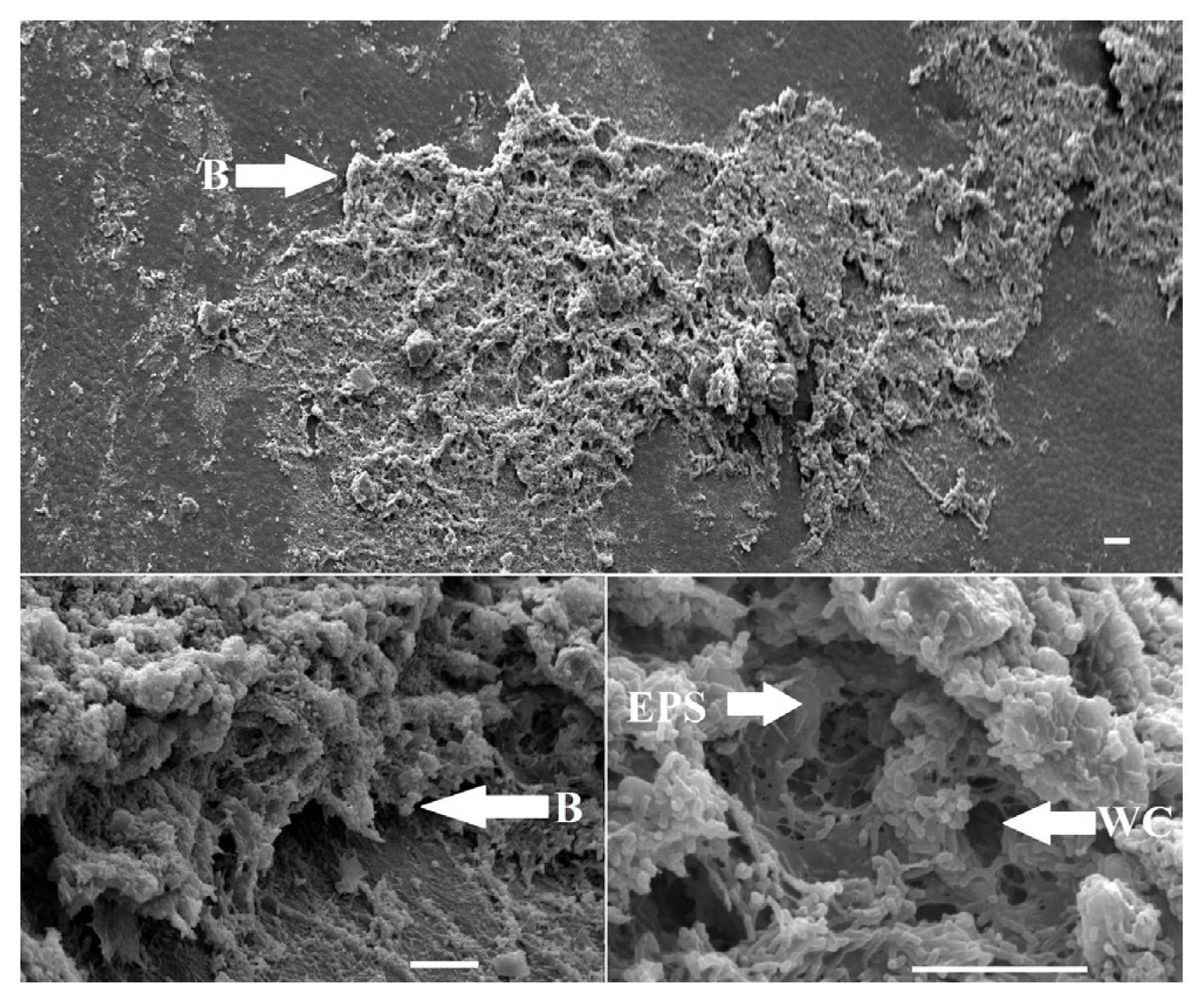
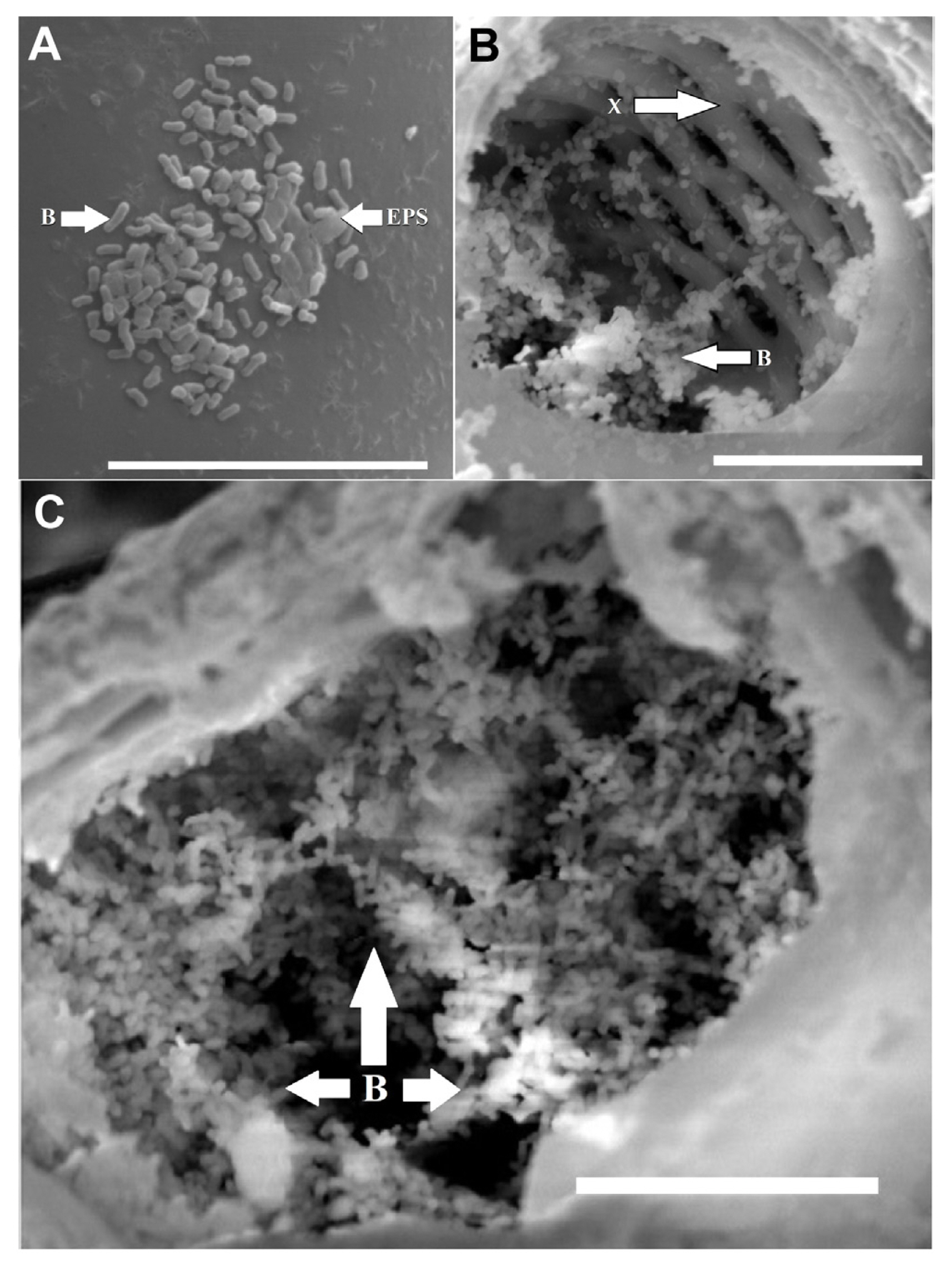
Bacterial growth in planta was visualized using SEM. Bacteria were clearly seen attached to the seed coat of discolored bean seeds. The bacteria were rod-shaped, and were not observed as solitary cells, but were dramatically clustered in biofilms (Fig. 4). Cells were clustered in multi-layer colonies and encased in what appeared to be the remnants of exopolymeric substances (EPS) (Fig. 4). Matrix-associated clustered layers of bacterial cells were not always densely packed, but had numerous openings and channels (Fig. 4). Surface-associated biofilm colonies were seen in categories 1 and 2, but were absent in categories 3 and 4 (not shown).
Leaf tissues from plants grown from category 2 seeds were also visualized via SEM. Occasionally, modest aggregates of small rod-shaped bacteria were seen on the surfaces of leaf tissues along with what appeared to be an EPS (Fig. 5). A number of internal vessels showed aggregates of rod-shaped bacteria within them (Fig. 5). Furthermore, colonization and occlusion of xylem elements with small rod-shaped bacteria within presumptive EPS was observed in category 2. Thus, evidence of biofilm, or pre-biofilm, formation by CFF cells was visible on seed surfaces, leaf surfaces and within vascular elements. No evidence of bacterial biofilms were observed in categories 3 and 4. Leaf tissues for category 1 could not be evaluated for biofilms on/in plants because emergence levels were zero in greenhouse pots.
Sensitivity of CFF biofilms and planktonic cells to bactericidal chemicals
The growth of CFF in the MBEC Assay reactor was evaluated and the bacterium was able to achieve 5 log10 colony forming units per mL (cfu/ml) within biofilms in 48 h. When challenged with bactericides, planktonic cultures were reduced to zero (i.e., total kill was achieved), with MBCs of less than 10 μg/ml (Fig. 6). ANOVA of the MBC values revealed no significant differences (P ≤ 0.05) between treatments (f-ratio value = 0.12059, and P = 0.974343). The counterpart CFF cells within biofilms were not so easily killed as MBEC curves consistently had higher numbers of survivors than MBC curves, and much higher concentrations were required to reduce the populations to zero or near zero (Fig. 6). For example, all MBC values were less than 10 μg/ml, but some MBEC values were more than 100 times greater. The bactericides with the largest gap between the MBC and MBEC curves were copper hydroxide (CuOH), oxysilver bisulfate (OSBS), and oxysilver nitrate (OSN). For these three bactericides, a Student’s t-test confirmed that the MBEC curve diverged significantly from the MBC curve between zero and 200 μg/ml (Table 2). ANOVA of the MBEC values revealed no significant differences (P ≤ 0.05) between treatments (f-ratio value = 0.30369, and P = 0.87378).
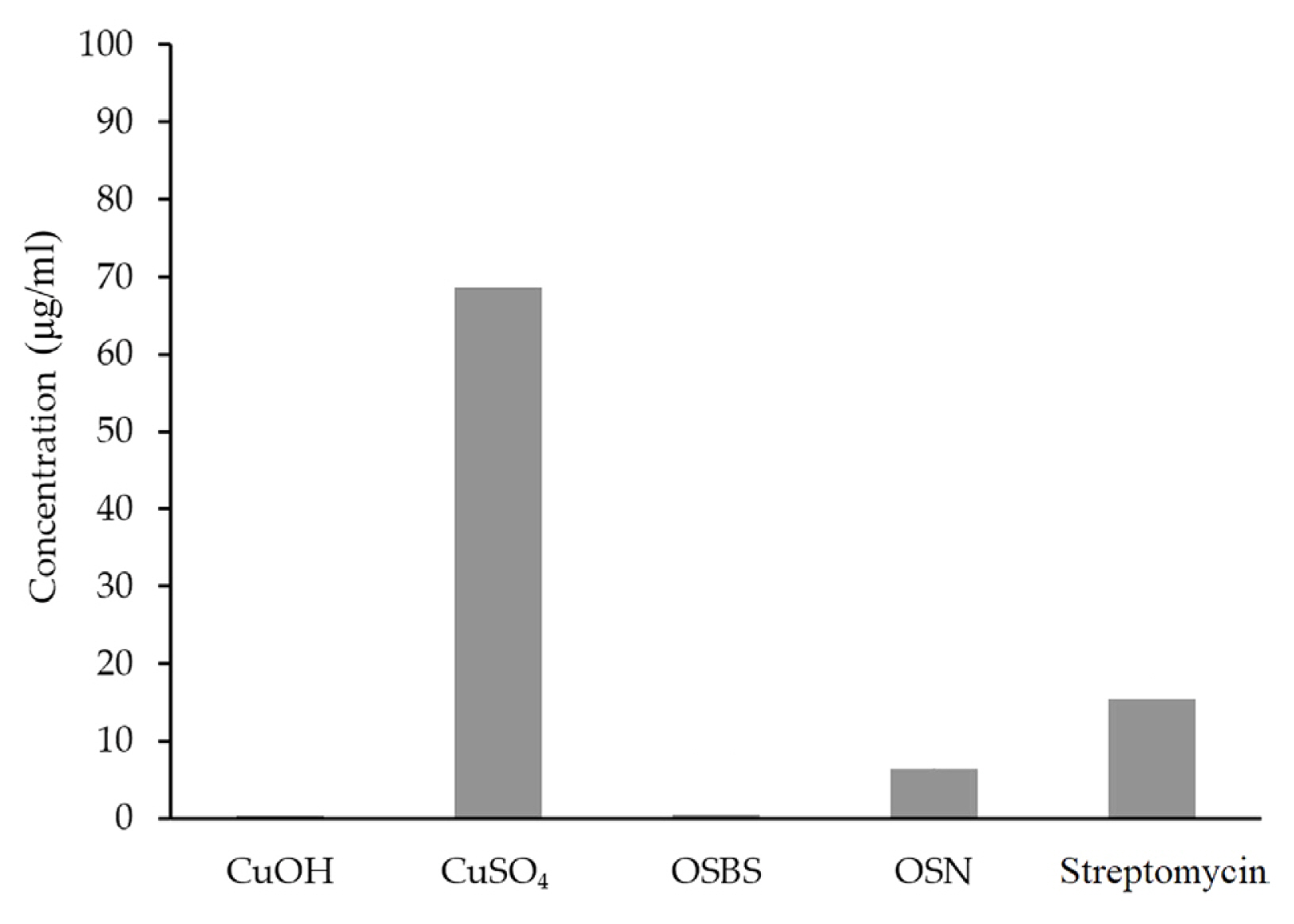
To further assess the bactericidal treatments’ efficacies, the EC50 was calculated for each. Copper hydroxide (Cu(OH)2) (0.32 μg/ml) and OSBS (0.49 μg/ml) had the lowest EC50 values followed by OSN (6.42 μg/ml). Streptomycin (15.51 μg/ml) and CuSO4 had the highest EC50 values (68.63 μg/ml), as seen in Fig. 7.
The EC50 results are valuable for comparing efficacies of the bactericides, however the portion of the curve being evaluated at 50% reduction is not very revealing when viewed from a practical perspective. For example, when dealing with bacterial wilt, one would like to reduce populations by ≥95%, because this is the level of reduction necessary to prevent disease spread. For this reason, the EC95 value was also calculated for each bactericide in order to evaluate the MBEC curves within the window that reductions in populations become more practically meaningful. The EC95 values show that OSBS (26.89 μg/ml) and streptomycin (95.35 μg/ml) were the most practically effective bactericides for reducing CFF biofilms by ≥95%, at the lowest concentrations, whereas the other three required concentrations from nearly 500 to over 1,300 μg/ml (Fig. 8).
Discussion
CFF is a re-emerging pathogen causing bacterial wilt on dry edible bean worldwide. It has multiple colony color phenotypes and is genotypically diverse (Agarkova et al., 2012). The bacterium grows systemically within bean plants, causing severe wilting symptoms and significant yield losses, and is seed-transmitted (Harveson and Schwartz, 2007). Highly effective chemical control options, such as seed- and foliar-applied bactericides are not currently available. The pathogen causes reductions in emergence (Fig. 3), and systemic biofilm infections that can lead to blockage and occlusion of the host vascular system (Fig. 5). These blockages can cause catastrophic dysfunction in water transport within leaves and serious yield and economic losses to producers.
Genetic resistance is often one of the best management tools for mitigation of diseases in crops. Some sources of wilt resistance have been characterized in bean (Conner et al., 2008; Hsieh et al., 2005; Urrea et al., 2008), but most or all of the dry bean cultivars grown in North America have not had wilt resistance introgressed or selected (Agarkova et al., 2012). This leaves clean seed programs and a few minimally effective seed treatments as the only management tools available for management of this disease.
It is thought that multiple factors likely combined to facilitate the re-emergence of this pathogen, with increased contaminated seed lots likely being a primary driver. CFF can form robust biofilms on seed that are exceedingly difficult to eliminate or eradicate (Fig. 4). Other factors driving the re-emergence of bacterial wilt may include climate change, enhanced adoption of center pivot irrigation, and increasing CFF survival on other crop hosts such as corn, soybean, wheat, and alfalfa (Agarkova et al., 2012).
Since contaminated seed lots are an important aspect of CFF resurgence, a biofilm approach to bactericide testing was taken, because biofilms are notorious for having much higher survival rates after antibiotic or biocide treatment, compared to planktonic, solitary or free-floating cells (Anwar et al., 1990; Bridier et al., 2011). Thus, since biofilms are the predominant growth form found in natural and agricultural settings, growing and testing of biofilm cultures in the lab is more likely to generate novel, efficacious field or clinical solutions (Harding et al., 2019; Villa et al., 2017). CFF was confirmed as another example of a plant pathogenic microorganism that forms biofilms during the disease cycle, as seen in the SEM images of bean seeds and within leaf veins (Figs. 4 and 5). The bacterium is present as multi-layered, surface-associated cells that appear to have been encased in a self-produced EPS. The openings and channels within the biofilm on seeds (Fig. 5) are characteristic of water channels commonly described in bacterial biofilms (Wilking et al., 2002, 2013). These channels facilitate convective transport and diffusion of solutes throughout the biofilm (De Beer et al., 1994, 1996; Picioreanu et al., 2000), which is essential for delivery of nutrients and removal of wastes. These morphological characteristics confirm that CFF is present within vessels, and on bean seed, as a biofilm.
In addition to the morphological appearance, CFF also displayed a hallmark phenotype associated with biofilms - increased survival after challenge with an antibiotic or biocide (Lewis, 2001). For example, in Fig. 6, the killing curves for planktonic cells drop to zero at low concentrations, but the biofilm cultures have a small subset of the population that survives at ≥100 times higher concentrations. Persister cells are a distinct subset of a microbial population that has temporary resistance or tolerance to conditions that are lethal to the majority of the population. Persister cells were described first in a Staphylococcus sp. (Bigger, 1944), and have since been characterized in many reports. The persister cell resistance phenotype is temporary, and distinguishable from antibiotic resistance arising as a result of genetic changes. Persister cells are much more common in biofilm populations compared with planktonic populations (Miyaue et al., 2018). Persister cells, along with other processes and phenomena, are likely responsible for the enhanced survival of biofilm populations treated with antibiotics and biocides (Stewart, 2015; Wood, 2017; Yan and Bassler, 2019), and may be responsible for the different killing curves for planktonic and biofilm cells seen in Fig. 6.
Since clean seed programs and seed treatments are the last line of defense against bacterial wilt, five bactericidal chemicals were compared for efficacy versus CFF biofilms. The results for EC50 showed that copper sulfate and oxysilver bisulfate were effective at the lowest concentrations (Fig. 7), but a 50% reduction in population is not sufficient to manage CFF infections on seeds. Therefore, the EC95 were also calculated and comparing them demonstrated that the bactericides effective at reducing the population by 95% at the lowest concentrations were streptomycin and oxysilver bisulfate (Fig. 8). Since streptomycin use as a seed treatment on dry bean is no longer supported by Health Canada, Canadian dry bean producers no longer have an approved efficacious bactericidal seed treatment option. Based on the results seen in Figs. 6,-8, oxysilver bisulfate appears to be a viable alternative to streptomycin. Streptomycin works against bacteria such as CFF by interrupting ribosomal functions. It binds irreversibly to the 16S rRNA located on the 30S component of the bacterial ribosome, inhibiting further protein synthesis by interfering with the assembly of initiation complex between mRNA and the bacterial ribosome. Additionally, Streptomycin leads to misreading of the mRNA and causes translational frameshift, thereby resulting in premature termination (Demirci et al., 2013). Silver and copper compounds’ mechanisms of action against microbes are based on their redox potential. These mechanisms of action include damaging of bacterial cell membranes, accumulation within the cells of ions that then react with and damage various cellular components, degradation of DNA and RNA, and respiratory chain inhibition (Bragg and Rainnie, 1974; Grass et al., 2011; Mathews et al., 2013; Modak and Fox, 1973; Russell and Hugo, 1994; Slawson et al., 1992). Thus, in general, silver is more effective than copper due to its higher redox potential. Oxysilver nitrate and oxysilver bisulfate are high oxidation state silver compounds, containing silver in the (I), (II), and (III) states, further increasing the efficacy relative to the copper compounds tested in this study, which were only present in the +1 state (Nadworny et al., 2015). A study comparing oxysilver nitrate and oxysilver bisulfate chemistry showed that oxysilver bisulfate is less stable and interacts more rapidly with water, which would result in more rapid release of antimicrobial species from the compound, which likely explains its higher efficacy (Nadworny et al., 2015).
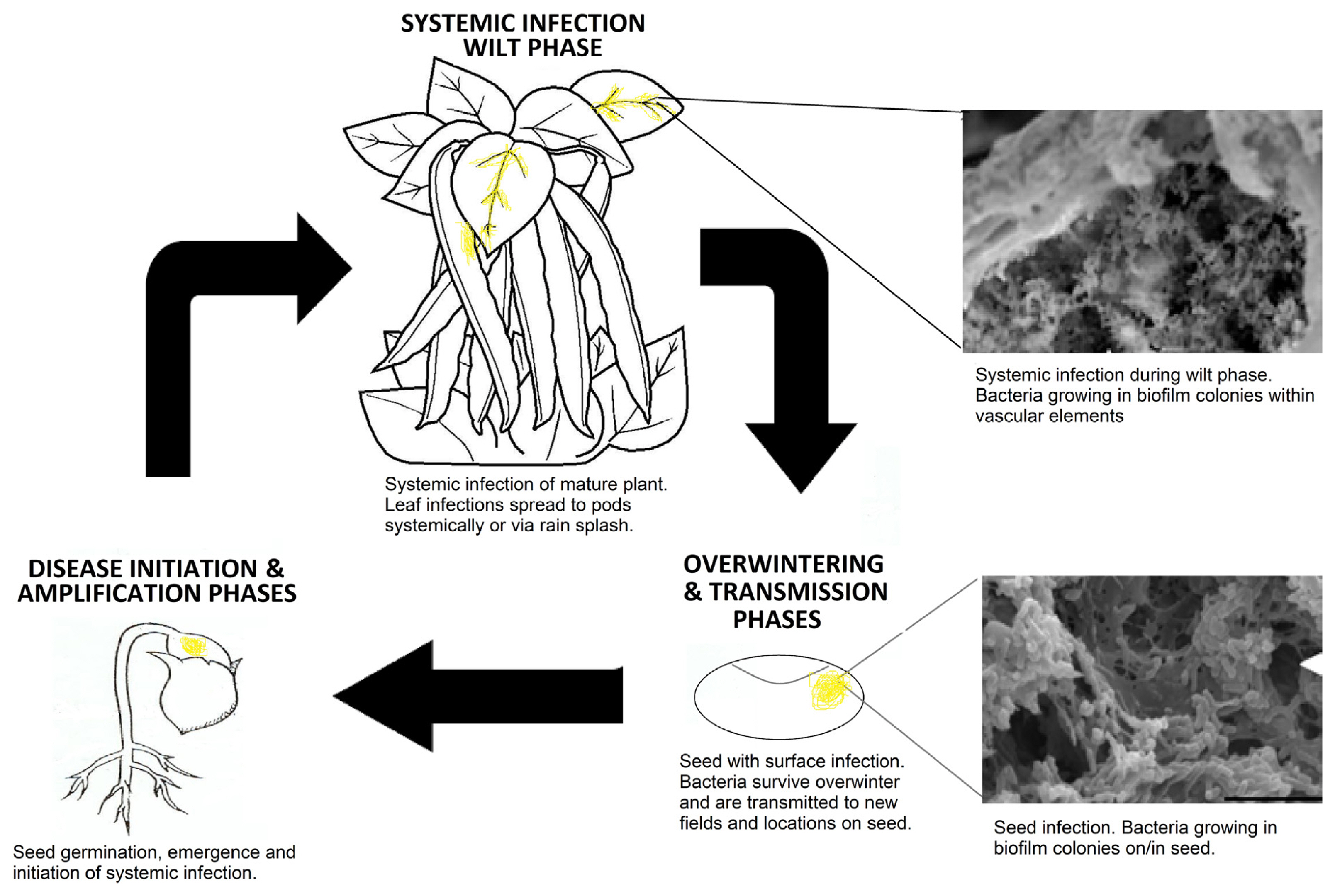
This is the first report in which a plant pathogen was demonstrated to form biofilms during two different phases of the disease cycle. In this case, CFF forms systemic biofilms within vascular elements of dry bean plants during the wilt phase of the disease, and then again on seeds as part of the overwintering and seed transmission phases (Fig. 9). The biofilm testing methodology described in this study demonstrates that CFF does form biofilms, and that these biofilm populations may play an important role in the transmission and resurgence of this pathogen on infected seed. This biofilm approach has also identified a bactericidal compound (oxysilver bisulfate) that may be a suitable replacement for streptomycin as a bactericidal seed treatment in areas where streptomycin use on seed is no longer supported by regulatory agencies.





 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






