



Fusarium head blight (FHB) is one of the most critical diseases of small-grain cereals such as wheat and barley. It causes not only damaged grains and yield loss, but also mycotoxin accumulation in infected grains. Fusarium asiaticum and F. graminearum, which belong to Fusarium graminearum species complex (FGSC), are two major contributors to FHB in these crops, particularly in Korea (Del Ponte et al., 2022; Jang et al., 2019; Lee et al., 1985; Shin et al., 2018). To date, five members (F. graminearum, F. asiaticum, F. vorosii, F. boothii, and F. meridionale) of the FGSC have been reported to be present in Korea with F. asiaticum predominant (Lee et al., 2016). These species can produce various mycotoxins including deoxynivalenol (DON) and its acetyl derivatives, nivalenol (NIV) and its acetyl derivative, zearalenone (ZEN), etc. (Desjardins, 2006; Lee et al., 2016). Unlike F. graminearum, which is widespread worldwide and mostly produces DON, F. asiaticum produces mostly NIV and is distributed with regional limits (van der Lee et al., 2015) and/or in relation to climate and cropping system (Lee et al., 2009; Xu et al., 2021).
In Korea, F. asiaticum with NIV chemotype is dominant in barley and wheat fields along with F. asiaticum and F. graminearum with DON chemotype. Barley and wheat are mostly cultivated consecutively with rice, allowing these crops to share FHB inocula for years and become conducive to infection. FHB severity can be managed by applying host resistance, chemical fungicides, biological control, or cultural practices (Buerstmayr et al., 2020; Chen et al., 2019). FHB resistance is an important agronomic traits for breeding barley and wheat (Steiner et al., 2017). However, few barley and wheat varieties exhibit strong and stable resistance in the fields (Bai and Shaner, 2004; Chen et al., 2019). Those developed for FHB resistance appeared to lose their resistance quickly over time, or many germplasms identified turned to variable and susceptible to FHB (Ma et al., 2020). In Korea, only a few cultivars are known to be resistant to FHB (Han and Kim, 2005; Kim et al., 2020; Ma et al., 2020; Park et al., 2008), and none of them impart satisfactory FHB or mycotoxin resistance.
In this study, to aid the selection of resources resistant to FHB, we evaluated barley and wheat germplasm accessions by testing FHB severity and mycotoxin accumulation. In the FHB assays, we used both F. asiaticum and F. graminearum as inocula to investigate whether they could produce similar levels of FHB in barley and wheat because FHB assessment for resistance is usually done with a single species. Here, we report the results of an evaluation of 60 germplasm accessions and a comparison of the FHB severity and mycotoxin production between F. asiaticum and F. graminearum.
Materials and Methods
Germplasm selection
Sixty germplasm accessions were selected from barley and wheat from the National Agrobiodiversity Center at the Rural Development Administration based on their agronomic traits. The selection was made among those with rapid heading, rapid ripening, and short stems. Additional traits were less cold damage and no lodging for barley, and short awns, high processing suitability, and resistance to FHB, powdery mildew, and leaf rust for wheat.
FHB assay
For the FHB assessment, the seeds of each accession were sown in peat soil and vernalized for 4 weeks at 5┬░C. The seedlings were transplanted into larger pots (25 cm in diameter) in a greenhouse. When the heads started to flower, five heads of each accession were inoculated by spraying (~2 ml/head) with conidia (1 ├Ś 105) of F. asiaticum #73 (Fa73) and F. graminearum Z3639 (Fg39) (McCormick and Alexander, 2002), as previously described (Baek et al., 2020). Fa73 was a strain isolated from Korean rice in 2015 and confirmed for NIV production. These strains were selected based on their ability to produce enough number of conidia and pathogenicity from our laboratory collection. Both strains were maintained at ŌłÆ80┬░C until use. Sterile water was used as negative control. The inoculated heads were covered with a plastic bag for humidity for three days and, then uncovered. Three weeks after inoculation, treated heads were harvested and scored for FHB severity. The assay was repeated twice during different seasons (March-May 2020 and November 2020-May 2021). FHB severity (%) was calculated as the mean of [(number of grains with FHB symptoms/total number of grains in a head) ├Ś 100]. Some heads that turned white after inoculation were excluded from FHB scoring.
Mycotoxin analysis
Harvested grains were completely dried in a hood and ground in a mixer mill (Retsch MM 400, Haan, Germany) for analysis of DON, NIV, 3-acetyl DON (3-ADON), 15-acetyl DON (15-ADON), or ZEN. Toxin analysis was performed as previously described with modifications (Lee et al., 2013). One gram of the ground sample was mixed with 8 ml of a solution of 1% acetic acid in acetonitrile (ACN) and extracted with shaking at 300 rpm for 1 h. After adding 0.25 g NaCl and 1 g MgSO4, the extract was centrifuged at 3,600 rpm for 10 min. The supernatant was mixed with 0.15 g MgSO4, 0.05 g endcapped C18, and 0.05 g primary secondary amine then centrifuged again under the same condition. The supernatant was dried under N2 gas and the residue was reconstituted with 20% ACN. For liquid chromatography-mass spectrometry (Waters e2695 separation module, Waters 3100 MS detector, Milford, MA, USA) analysis, a Zorbax SB A1 C18 (4.6 ├Ś 150 mm, 5 ╬╝m, Agilent, Palo Alto, CA, USA) column was used. The flow solvent was a water mixture containing 0.2% formic acid and 5 mM ammonium formate in DW (A) and methanol (B) in a gradient. The mass spectrometer was operated in single ion recording (SIR mode), both in electrospray ionization positive and negative. The mobile phase flow rate was 0.3 ml/min and the injection volume was 10 ╬╝l.
Results
FHB severity
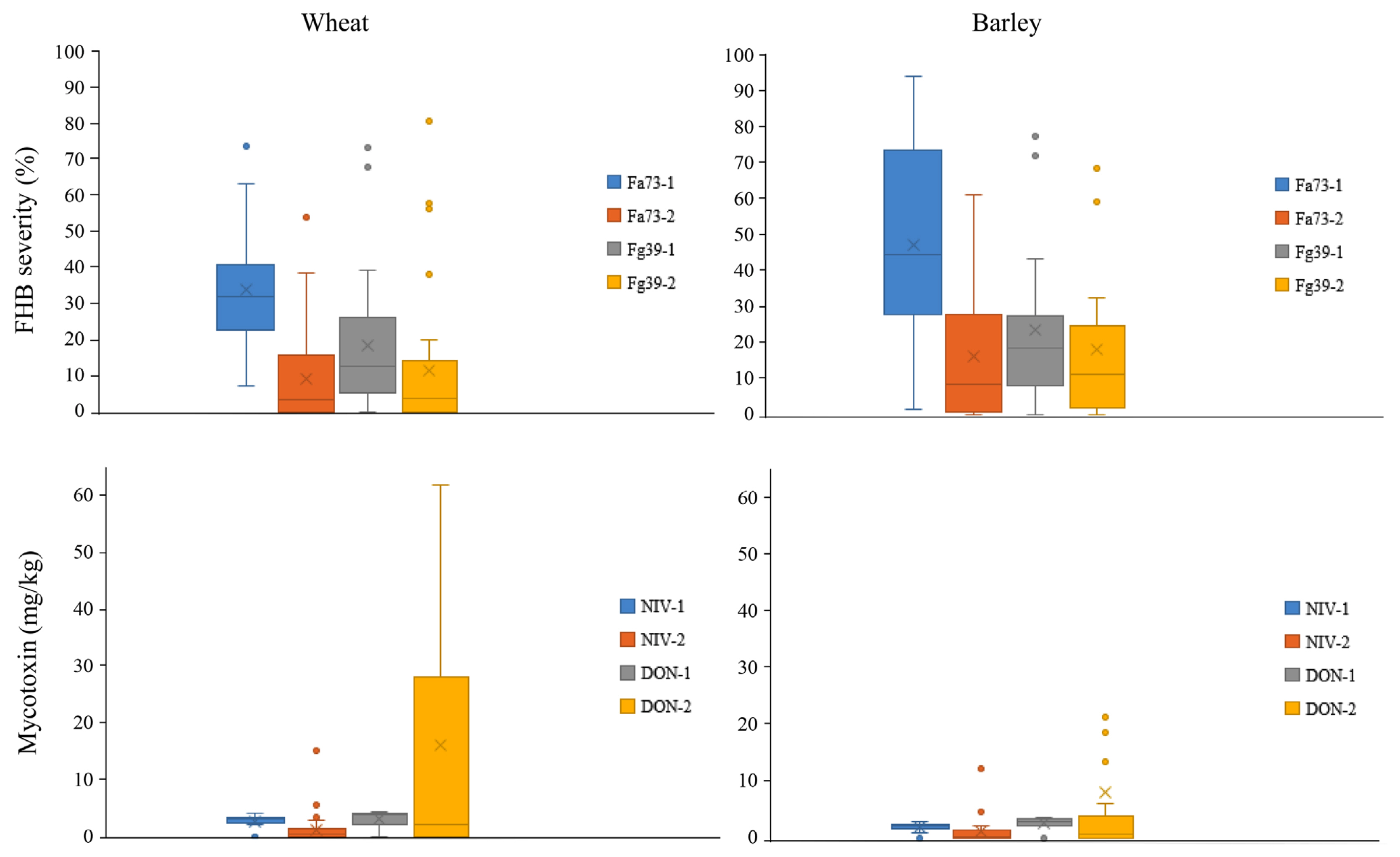
The germplasm accessions tested varied in FHB severity by inoculum or assay. However, FHB severity induced by either strain was greater in barley than in wheat. Fig. 1 shows clearly that the blue box (the result of the first assay with Fa73) in barley panel was greater than one in wheat by the values of mean, median, and maximum severity. Other colors show the same pattern. In the first assay, the means of FHB severity by Fa73 were 46.8% and 33.9%, ranging up to 93.6% and 73.6% in barley and wheat, respectively (Tables 1 and 2). The means of FHB severity by Fg39 were 22.7% and 18.3%, and up to 77.1% and 73.5% in barley and wheat, respectively. In the second assay, both strains produced much lower means of FHB severity than in the first, and Fg39 produced slightly higher means (18.0% in barley and 11.7% in wheat) than Fa73 (16.2% in barley and 9.3% in wheat). The inoculated heads of some accessions (especially 33% of barley) turned white in the second assay, showing the sensitivity of this assay (Table 1).
The lowest FHB severity was found in one barley (b32) (mean 10.5%, ranging from 8.0% to 15.3%) (Table 1). In wheat, the lowest were two (w49, w51) with means of 11.4% and 4.6%, ranging from 0.9% to 18.2% and from 0 to 18.2%, respectively (Table 2). There were two more wheat accessions with the highest FHB severity of 19.0% (w3) and 19.5% (w12) (Table 2). The rest showed >20% FHB severity. Among the 19 barley accessions with severity values, 18 (95%) showed higher severity by Fa73 than by Fg39 in the first assay, and 14 of them (74%) showed >30% FHB severity by Fa73 (Table 1). Only four of them showed >30% FHB severity by Fg39. In the second assay, six accessions were consistent with higher severity by Fa73 (Table 1), whereas six others turned to the opposite with higher severity by Fg39. Only two and three accessions showed >30% FHB severity by Fa73 and Fg39, respectively. One accession (b60) was consistent with a higher severity of Fg39. In wheat, 28 (85%) of 33 accessions with severity values showed higher severity for Fa73 than for Fg39 in the first assay (Table 2). Nineteen of the 33 accessions (58%) showed >30% FHB severity by Fa73, and four of them had >30% FHB severity by Fg39. In the second assay, 10 accessions maintained higher severity by Fa73, whereas 12 others turned to the opposite with higher severity by Fg39. Only two and four accessions showed >30% FHB severity by Fa73 and Fg39, respectively. None of the wheat accessions were consistently susceptible to Fg39.
Mycotoxin production
Both NIV and DON were detected in most of the accessions analyzed with greater mycotoxin accumulation by Fg39 (Fig. 1). In the first assay, Fa73 produced means of 2.1 and 3.1 mg/kg NIV in barley and wheat, respectively, ranging from 1.1 to 3.0 mg/kg in barley, and from 0.3 to 4.0 mg/kg in wheat (Tables 1 and 2). Fg39 produced means of 3.0 and 3.7 mg/kg DON in barley and wheat, respectively, ranging from 0.3 to 3.7 mg/kg in barley, and up to 4.4 mg/kg in wheat. In the second assay, Fa73 produced decreased level of NIV than the first but with increase in some accessions up to 12.4 mg/kg (barley) and 15.1 mg/kg NIV (wheat) (means of 1.3 mg/kg and 1.2 mg/kg in barley and wheat). In contrast, Fg39 produced increased level of DON up to 120.5 mg/kg and 97.4 mg/kg in barley and wheat, respectively (means of 9.0 mg/kg and 16.2 mg/kg in barley and wheat). However, all medians were much lower than the first. It is notable that the highest DON and NIV levels were detected from the very same accession with >60% FHB severity (b41) (Table 1), indicating the susceptibility of this accession especially to mycotoxin accumulation. Regardless of the FHB severity of the accessions, the level of DON detected was higher than that of NIV in most accessions. Additional analysis of the acetyl derivatives of DON in the second assay showed that both 3-ADON and 15-ADON were produced in 29% and 75% of barley, and in 42% and 61% of wheat, respectively. Between 3-ADON and 15-ADON, the levels of 15-ADON were higher than 3-ADON levels in all accessions with one exception. The accessions with >10 mg/kg DON also accumulated 15-ADON up to 33.8 mg/kg, ranging in sum production of DON and acetyl DONs from 12.7 to 132.3 mg/kg. The five accessions with <20% FHB severity accumulated either NIV or DON up to 3.9 mg/kg at maximum, except one (w49) with accumulation of 41.1 mg/kg of DON and 10.7 mg/kg of 15-ADON. ZEN was detected in only 8% of barley (2/22), with mean of 0.1 mg/kg (Table 1).
Correlation between FHB severity and mycotoxin levels in barley and wheat
To investigate the associations between FHB severity and toxin level by F. asiaticum and F. graminearum, correlation was analyzed between disease severities by both strains in each crop, toxin levels by both strains in each crop, and between disease severity and toxin level by each strain in each crop. Firstly, FHB severities between two strains (Fa73 and Fg39) were moderately correlated in both crops (r = 0.57 and 0.60 in barley, 0.42 and 0.58 in wheat) (P < 0.05) (Table 3). Second, mycotoxins between the two strains (NIV by Fa73 and DON by Fg39) were highly correlated in barley (r = 0.69 and 0.91), but not in wheat (r = ŌłÆ0.08 and 0.06). Third, FHB severity by either strain was correlated with mycotoxin levels, especially in barley. In barley, the correlation between FHB and NIV was moderately high (r = 0.42 and 0.82), whereas that between FHB and DON was less consistent (r = 0.22 and 0.76). In wheat, however, the correlations were high only in the second assay (r = 0.76 by Fa73, 0.66 by Fg39). Due to the different numbers of datasets, disease severity or mycotoxin levels between wheat and barley were not analyzed.
Discussion
Germplasm accessions showed diversity in FHB resistance and mycotoxin accumulation. Most accessions (>91%) showed >20% FHB severity caused by either F. asiaticum or F. graminearum, and none of the accessions were free from mycotoxin accumulation. Among the five accessions with <20% FHB severity, barley (b32) was associated with the lowest level of mycotoxin (up to 2.2 mg/kg), suggesting the possibility of double FHB resistance (type II-resistance to spread and type III-resistance to toxin accumulation) (Chen et al., 2019). These results demonstrate that such FHB resistance can be secured and that FHB resistance is better regarded not as a single disease resistance but as a double resistance to FHB and mycotoxins. The resistant accessions revealed in this study can be useful resources for breeders. Both FHB and mycotoxin production are significantly affected by environmental and experimental conditions. Such effects of plant conditions and micro-environments can cause inconsistent evaluations. Although we tried to keep the assay conditions such as inoculum concentration, pathogen viability and plant conditions consistent, there could be variations in the temperature and humidity of the greenhouse between experiments as the accessions were tested in several months during a season. Thus, higher FHB severity should be counted, even with discrepancies between assays, because they reflect the susceptibility of a tested variety or accession under given environments.
Comparison of FHB severity by both F. asiaticum and F. graminearum was conducted in the germplasm for the first time in this study. The barley and wheat accessions showed different responses to F. asiaticum and F. graminearum under the test conditions. Our results suggest to use both species for FHB assessment in barley and wheat in Korea. FHB severity was generally higher in barley than in wheat in both strains, showing that barley was more susceptible to FHB, confirming our previous results (Baek et al., 2020). Of these two species, NIV-producing F. asiaticum was more aggressive than DON-producing F. graminearum in both crops. However, the latter had a tendency to be more aggressive toward wheat. Although the pathogenicity of Fa73 appeared to be higher than that of Fg39, the absolute amount of mycotoxins produced was greater by Fg39. The mycotoxin levels were higher in wheat for both DON and NIV, indicating that wheat was more prone to mycotoxin accumulation. These results differ from those of a previous study where DON levels were not significantly different between barley and wheat, and DON-producing F. graminearum was less aggressive than NIV-producing F. asiaticum in wheat (Baek et al., 2020). This work, however, was based on a lower number (14 in total) of cultivars, making our results more representative. Other studies reported that NIV-producing F. asiaticum and DON-producing F. graminearum were not significantly different on virulence in wheat cultivars (Jang et al., 2019; Shin et al., 2018). However, these results were based on a single cultivar or on a single pathogenicity test, respectively. Notably, DON production is accompanied by acetyl derivatives such as 3-ADON and 15-ADON. The toxicity of these acetyl DON derivatives is less than that of DON, but they still contribute to total toxicity (Knutsen et al., 2017). However, little information is available on the occurrence of acetyl DON derivatives in Korean cereals. Although Korean cereals are prevalent in NIV-producing F. asiaticum, they remain contaminated with DON, indicating that acetyl DON derivatives co-occur, as shown in this study. For food safety purposes, it is necessary to monitor DON derivatives in cereals and cereal-based products. Differences in FHB severity between F. asiaticum and F. graminearum and the superiority of F. graminearum in mycotoxin production (Gale et al., 2011; Jang et al., 2019) should be considered, especially for wheat breeding, as this crop is challenged more by F. graminearum. As wheat cultivation is currently encouraged in Korea to increase wheat self-sufficiency (Kim et al., 2020), it is important to secure wheat breeding resources that are resistant to both FHB and mycotoxins.
Our analyses confirmed previous results that FHB severity and toxin production are correlated in wheat and barley (Choo et al., 2004; Geddes et al., 2008; He et al., 2015; Yan et al., 2022). Unlike the previous studies, we used both F. asiaticum and F. graminearum at the same time and were able to compare these species. The highest levels of DON and NIV were associated with high FHB severity in both crops, especially F. graminearum (accessions b41 and w54). However, it is controversial to conclude because some studies found no correlation between FHB and DON concentration in these crops (Gilbert et al., 2002; Khanal et al., 2021) and all test conditions were different. The strong correlation between DON and NIV levels only in barley, and higher correlation between FHB severity and toxin production with F. asiaticum in barley accessions than wheat or with F. graminearum might be another proof of the co-evolution of F. asiaticum and barley for a longer period than wheat, as speculated previously (Baek et al., 2020). Further studies are necessary to elucidate the basis of this association between barley and F. asiaticum.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






