


Pectobacterium (previously classified as the genus Erwinia) belonging to the family Pectobacteriaceae are robust phytopathogens that cause soft-rot disease in economically important vegetables, fruits, and ornamental plants (Ma et al., 2007). Currently, this group has been classified into 20 species (P. carotovorum, P. odoriferum, P. versatile, P. brasiliense, P. atrosepticum, P. aroidearum, P. betavasculorum, P. fontis, P. cacticida, P. wasabiae, P. zantedeschiae, P. parmentieri, P. peruviense, P. polaris, P. punjabense, P. polonicum, P. aquaticum, P. actinidiae, P. parvum, and P. quasiaquaticum) based on advanced genomic analysis techniques (Jee et al., 2020; Moussa et al., 2021; Pasanen et al., 2020; Portier et al., 2019, 2020; Zhang et al., 2016).The genus is a gram negative, primarily rod-shaped bacteria and most of them are plant pathogens. Pectobacterium usually invades through wounds on the host plant caused by various factors like insects, other diseases, and physical factors (Oskiera et al., 2017). They can produce pectin and cellulolytic enzymes that break down cell walls, causing maceration (Charkowski, 2018). This series of processes can be repeated for a long time not only in the field but also in the storage and distribution process (Bhat et al., 2010; Glasner et al., 2008).
Kimchi cabbage (Brassica rapa var. pekinensis) is one of the economically significant stored vegetables produced and supplied year-round in Korea (Lee et al., 2016). However, it is vulnerable to various diseases and pests when exposed to high temperatures. In particular, soft rot, which occurs frequently occurs in Kimchi cabbage growing at high altitudes in summer, causes huge losses (Kim and Yeoung, 2004). Although P. carotovorum had been known to cause most Kimchi cabbage soft rot, P. odoriferum, P. brasiliense, and P. versatile were recently isolated from soft rot in Kimchi cabbage (Lee et al., 2014; Park et al., 1999; Roh et al., 2009; Seo et al., 2004). P. odoriferum was primarily isolated from chicory in France and Japan, and was initially recognized a narrow host range (Gallois et al., 1992). However, it has been isolated from various vegetables (celery, cabbage, Chinese cabbage, artichoke, carrot, parsley, onion, potato, sugar beet, and sweet potato), and has been identified as the causal agent of bacterial soft rot in many other countries (Gao et al., 2016; Oskiera et al., 2017; Waleron et al., 2014). For that reason, there is a need for rapid and accurate identification of pathogens for an appropriate disease control strategy for important stored vegetables, including Kimchi cabbage.
Identification of P. odoriferum has traditionally been based on morphology and host range, but most hosts overlap, and the phenotypic characteristics are unclear depending on growth conditions (Czajkowski et al., 2015). These traditional methods are poorly reproducible, time-consuming, and labor-intensive. Over the past three decades, ribosomal RNA operons, specifically the 16S rRNA genes, have been widely used for the diversity analysis and identification of prokaryotes. However, sequencing of the 16S rRNA gene has not differentiated the species of the genus Pectobacterium (Naum et al., 2008; Staley, 2006). Although some molecular methods have been developed for rapid detection and identification, using molecular methods such as amplified fragment length polymorphism and multi-coordinate sequencing (Nabhan et al., 2012). Iterative and time-consuming procedural difficulties usually accompany this species identification method. Subsequently, a phylogenetic analysis method was developed based on the single gene pmrA required for resistance to plant-derived antimicrobial peptides (Kettani-Halabi et al., 2013). The tests must be rapid, sensitive, and specific, as cost- and time-sensitive decisions are made when implementing a disease control strategy. In addition, two specific primers for P. odoriferum are six genes of the Pco-subspecific srl operon (i.e., srlAEBDMR) involved in sorbitol metabolism. Two specific primers for general polymerase chain reaction (PCR) and quantitative PCR (qPCR) based have been developed (Li et al., 2020).
We designed species-specific primers for general PCR and qPCR in this study based on the HAD family hydrolase gene. In addition, specificity and sensitivity were tested for the detection of pathogens in infected cabbage leaves according to the MIQE guidelines (Bustin, 2008). Currently, real-time qPCR is known as a rapid and accurate approach for pathogen diagnosis in plants. Therefore, we proposed an efficient method for specific and quantitative detection of the pathogen, P. odoriferum, in plant samples through direct qPCR based on SYBR Green. Thus, it shows that it can be used for sensitive and specific detection of target pathogens, simplifying diagnosis, pathogen monitoring and implementing of appropriate disease control strategies.
Materials and Methods
Bacterial culture and genomic DNA isolation
Twenty-nine culture were selected for the experiment. Among them, 24 type strains were provided from the Korean Agricultural Culture Collection (KACC) and five strains of P. odoriferum isolated from the Republic of Korea obtained from KACC and the Highland Agriculture Research Institute to validate the specificity of the species-specific primers developed in this study (Table 1).
The bacterial isolates were cultured at 28-30┬░C for 24-48 h on trypsin soy agar media (15 g/l digested casein, 5 g/l digested soy, 5 g/l NaCl, and 1.5% agar, Difco, Detroit, MI, USA) under aerophilic condition. The stock culture were preserved in glycerol stocks at ŌłÆ80┬░C. Total genomic DNA was extracted using the NucleoSpin Microbial DNA Kit (TaKaRa Bio USA, Inc., San Jose, CA, USA). The corresponding strains listed in Table 1 and isolated DNA was spectrophotometrically quantified. The concentration and quality of the extracted genomic DNA were determined using a NanoDrop ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). All the extracted genomic DNA samples were stored at ŌłÆ20┬░C until used for further experiments.
Gene selection and oligonucleotide primer design for species-specific PCR assay
The whole genome sequences from P. odoriferum JK2.1 (GenBank accession no. CP034938.1) and the other closely related species data were downloaded from the NCBI ftp site (ftp://ftp.ncbi.nlm.nih.gov/genomes/bacteria/). To mine species-specific genes for quantitative detection of the targeted microbe, candidate gene selection pipeline via computational clustering was carried out as described by Chen and Lang (Chen et al., 2010; Lang et al., 2010). From this candidate gene selection pipeline, the resulting candidate genes presented no substantial concordance with other P. carotovorum group strains that were selected as quantitative PCR targets. The specific primer set used for P. odoriferum was generated using the DNASTAR Lasergene Primerselect module (version 7.2.1, DNASTAR Inc., Madison, WI, USA) (Table 2). Primer synthesis was performed by XENOTECH Co. (Daejeon, Korea). The oligonucleotide primer set amplified a specific DNA fragment from only the targeted species. The nucleotide sequences of the primer set were evaluated for their specificity via NCBI BLAST modules such as BLASTn and BLASTx (https://blat.ncbi.nlm.nih.gov/Blast.cgi).
Conventional PCR protocol
PCRs were conducted in T100 Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA) with the following steps: initial denaturation at 95┬░C for 5 min; 35 cycles of denaturation (at 95┬░C for 1 min), annealing (at 65┬░C for 30 s), and extension (at 72┬░C for 1 min), and a final extension period of at 72┬░C for 10 min. All PCR reactions were performed in a total volume of 25 ╬╝l (1├Ś buffer, 0.2 mM each dNTP, 4.0 mM MgCl2) covering 1.25 U of GoTaq Flexi DNA polymerase (Promega, Madison, WI, USA), 25 ng of template DNA and a 0.1 ╬╝M final concentration of each primer (Table 2). PCR product was resolved by 1.5% (w/v) agarose gels and stained with LoadingStar (DYNEBIO, Seoul, Korea). The gel was visualized and the images were documented via a GelDoc Go Imaging System (Bio-Rad Laboratories).
SYBR Green quantitative PCR conditions
All quantitative PCR assays were carried out in a final reaction volume of 20 ╬╝l containing 5 ng of purified DNA extracted from each sample, primers mentioned above (0.5 ╬╝M final concentration) and QGreenBlue 2├Ś Green qPCR Master Mix (CellSafe Bio, Inc., Yongin, Korea). The SYBR Green qPCRs were performed in a CFX96 real-time PCR system (Bio-Rad Laboratories). The SYBR Green real-time PCR amplification conditions were as follows: initial denaturation of at 95┬░C for 30 s; 40 cycles at 95┬░C for 5 s, 65┬░C for 30 s, and a melting curve analysis from 65 to 95┬░C with an increment of 0.5┬░C per 5 s. The standard curve of P. odoriferum was created by plotting the qPCR cycle threshold (Ct) values conducted using 10-fold serial dilutions of genomic DNA, cloned DNA (pGEM-T easy vector containing the 360 bp species-specific amplicon) and a bacterial cell suspension of targeted microbe. The copy number of the template was calculated using the previously described formula (Kang et al., 2016; Whelan et al., 2003):
The Bio-Rad CFX Maestro version 1.1 software suite automatically performed thermal cycling, data collection, and standard amplification. A non-template negative control and standard amplification were also included to confirm the SYBR Green real-time PCR quality. The amplification efficiency (E) was calculated using the following formula.
Pathogen detection and quantification by real-time quantitative PCR
In order to detect and quantify pathogen in Kimchi cabbage leaves infected with P. odoriferum, the bacterial cell suspension (2.47 ├Ś 107 cfu/ml, 50 ╬╝l/inoculation site) was injected in two places on the lower leaf veins of four healthy Kimchi cabbage leaves. After three days of post-inoculation, soft root disease symptoms were confirmed, and samples were collected at five locations (A-E, 20 mm interval) by distance along the leaf vein from the site of disease progression. Each sample (leaf pieces about 1 ├Ś 1 cm) was collected using a sterilized instrument and immersed in a 2 ml tube containing 1 ml of sterile water for about 30 min, and 1 ╬╝l of the immersion solution was used for real-time qPCR. The limit of detection (LOD) corresponds to undetected amplification when Ct Ōēź 37, and the limit of quantitation (LOQ) corresponds to a Ct value when the threshold standard is not met or the standard deviation value is 1.0 Ct Ōēź 36 that not meet the threshold standard in 3 replicates or a standard deviation value of 1.0 or greater. For quantitative analysis, the density of pathogens by distance from symptom was estimated by referring to the cell suspension standard curve results.
Results
In silico specificity assay of the selected genes and primer design
The selected gene and oligonucleotide primer set (Table 2) from P. odoriferum were evaluated and confirmed via a combination of Bioinformatics tools, including the Lasergene PreimerSelet program (version 7.2.1, DNASTAR Inc.) and NCBI BLAST search engines (http://blast.ncbi.nlm.nih.gov/cgi).
For a HAD family hydrolase-coding gene (accession no. CP034938.1, 3809023-3809706, protein ID: QHP81543.1) of P. odoriferum JK2.1, the BLASTn searches presented no substantial match to the recognized reference sequences of other Pectobacterium species. The BLASTx results using the predicted protein sequence presented that the most relevant protein to our HAD family hydrolase gene was a P. carotovorum subsp. carotovorum 2-deoxyglucose-6-phosphatase (identity = 99.12%, score = 468 bits [1,205], and expected = 1e-166).
Conventional and real-time PCR for verification of primer specificity
Genomic DNA samples from Pectobacterium, Dickeya, Pantoea, Erwinia, and Musicola strains covering the type strains of each targeted species were used to validate the specificity of the microbe-targeting oligonucleotide primer set via a conventional PCR test.
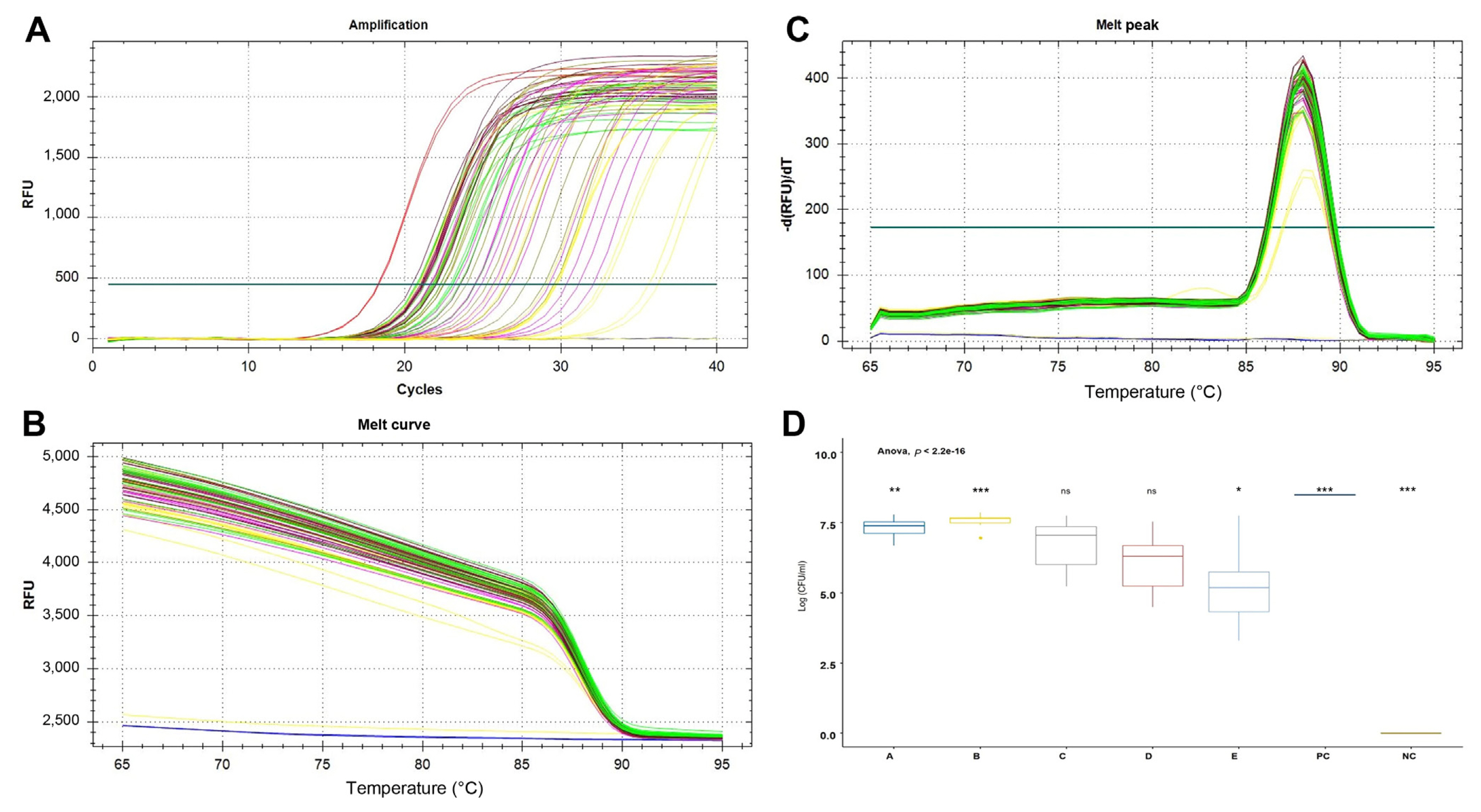
The specificity of the PeOd360F/R primer set was tested by conventional PCR using 29 different bacterial genomic DNA (Table 1). The target gene, expected size of 360 bp, was amplified only in the P. odoriferum strains (Fig. 1). In addition, SYBR Green-based real-time PCR correctly generated PeOd360F/R primer pair-specific fluorescence signals in all P. odoriferum strains (Fig. 2). On the other hand, other bacteria species had no fluorescence signal (Fig. 2, lanes 7-13 and 15-28). In addition, the melting peak showed reproducible single peaks at 88┬░C.
SYRB Green qPCR assay efficiency, LOD, and LOQ
SYBR Green-based real-time PCR was conducted with PeOd360F/R primers to generate a standard curve by plotting the mean threshold cycle (Ct) value (n = 3). The concentration of genomic DNA, cloned DNA, and density of the bacterial cells were 5 ├Ś 10ŌłÆ15 to 5 ├Ś 10ŌłÆ9 g/╬╝l, 1.35 ├Ś 102 to 1.35 ├Ś 109 copies/╬╝l and 2.47 ├Ś 101 to 2.47 ├Ś 107 cfu/ml, respectively (Table 3). The standard curves showed that there are linear-correlation between the Ct values and the concentration of genomic DNA (R2 = 0.999, slope = ŌłÆ3.688, and y-int = 38.709), cloned DNA (R2 = 0.999, slope = ŌłÆ3.928, and y-int = 29.093), and cell suspension (R2 = 0.998, slope = ŌłÆ3.995, and y-int = 44.407). The efficiencies were 86.7%, 79.7%, and 78.0% for each genomic DNA, cloned DNA, and cell suspension (Fig. 3). The LOD results showed that the fluorescence signal could be detected when the genomic DNA and cell suspension were diluted to 50 fg and 2.47 ├Ś 103 cfu/ml, but could not be detected when diluted to 5 fg and 2.47 ├Ś 102 cfu/ml. Therefore, the LOQ of cloned DNA by real-time PCR was confirmed to be 1.35 ├Ś 102 copies. The LOQ of genomic DNA and cell suspension by real-time PCR was confirmed to be 50 fg and 2.47 ├Ś 104 cfu/ml, respectively (Table 3).
Pathogen detection in leaf samples using SYBR Green-based real-time PCR
Direct detection and quantitation of the PeOd360F/R primer set were confirmed by SYBR Green real-time PCR in disease-symptomatic Kimchi cabbage leaf samples. The sample was divided by distance (A-E) from the symptom site and repeated 3 times.
As a result, the Ct values of region ŌĆ£AŌĆØ near the symptom site and region ŌĆ£BŌĆØ close to it were 22.30 ┬▒ 1.14 and 21.46 ┬▒ 0.78, respectively, significantly lower than at other locations somewhat distant from the symptom site. The positive control had a Ct value of 18.29 ┬▒ 0.04 and the negative control had no signal (Fig. 4). In addition, the number of cells was calculated based on the cell suspension standard curve. As a result, region A-E were confirmed to be 7.32 ┬▒ 0.33, 7.57 ┬▒ 0.23, 6.69 ┬▒ 0.88, 6.06 ┬▒ 0.96, and 5.14 ┬▒ 1.29 Log cfu/ml, respectively (Fig. 4D).
Discussion
The strains P. odoriferum, P. carotovorum, P. brasilience, and P. versatile is currently recognized as the primary the causal agent of bacterial soft rot on storage vegetables in the Republic of Korea (Lee et al., 2014; Park et al., 1999; Roh et al., 2009; Seo et al., 2004). However, the species do not have host specificity (P├®rombelon and Kelman, 1980). Therefore, soft rot disease diagnoses and identifying specific pathogens are economical essential. Furthermore, the microbe genome sequences and genomic analysis techniques can accelerated identification and disease diagnosis. As result, it provides an extended possibility to develop existing detection and quantitation methods by identifying novel target genes.
The methods adopted for rapid and accurate diagnosis include real-time qPCR analysis using SYBR Green-based and FAM-labeled TaqMan probe methods. The specific primer set for qPCR based on SYBR Green in the 16S-23S rRNA internal transcribed spacer (ITS) region was developed to detect and quantify P. carotovorum (Su├Īrez et al., 2022). In addition, the P. brasiliense-specific TaqMan probe-based qPCR assay for detecting and quantifying has been developed by comparing the 16S-23S rRNA ITS and the tRNA-Glu gene region which contain polymorphisms informative for the discrimination of closely related Pectobacterium species (Muzhinji et al., 2020). The specific primers for diagnosing P. odoriferum are currently the only primer sets for conventional PCR and SYBR Green-based qPCR reported by Li et al. (2020). They developed two primer sets based on the srlE gene, encoding sorbitol-specific phosphotransferase, to identify P. odoriferum (Li et al., 2020). However, their study did not show accurate pathogen quantitation values in infected plants. In addition, it requires some effort because DNA is extracted and used from infected plants.
In the present study, we used BLAST search to explore for P. odoriferum-specific genes. In addition, we developed the specific primer set based on the HAD family hydrolase gene involved in various cellular processes ranging from amino acid biosynthesis to translation (Koonin and Tatusov, 1994) (Table 2). The advantage of our newly developed assay is that it does not require pre-incubation and DNA extraction for species identification and quantification. In addition, accurate identification of P. odoriferum using conventional PCR and qPCR methods is possible with one primer set. The result of the calculated standard curve shows a small standard error. It ensures that the assay is reproducible and very robust, not only on pathogen gDNA but also on infectious plant suspensions. As a result of measuring the pathogen concentration by distance from the symptom, it was confirmed that the pathogen moved along the leaf veins, and when the infection progressed, the pathogen could be detected even in an area with no symptoms (Fig. 4). The LOD of pathogens is 103 cell units, enabling precise diagnosis.
In conclusion, the newly developed SYBR Green real-time qPCR primer set can perform qPCR directly from plants without DNA extraction. This method is susceptible, saving time, significantly reducing the risk of contamination and cost per reaction, and helping process processing large volumes of samples. The newly developed specific primer set can help in epidemiological study including where this pathogen present by early detection of P. odoriferum.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






