



Rice is the most widely consumed crop by more than half of the world’s population, particularly in Asia and Africa, and is produced in approximately 120 countries worldwide. Rice is the world’s fourth-most produced agricultural crop after sugarcane, maize, and wheat. Global production amounts to 756 million tons, of which 4.7 million tons is produced by South Korea, the 18th largest global rice producer (Food and Agriculture Organization of the United Nations, 2020). Rice is a staple crop in Korea with socio-economic significance. Fifty-eight percent (635,000 households) of all farming households in the agriculture and forest industry of the country are engaged in rice farming, accounting for the highest percentage in the industry (Jeong et al., 2017).
The name “bakanae” originates from the Japanese term “bakanae” that translates to “foolish seedling.” It is one of the oldest diseases reported in rice (Hori, 1898). Bakanae occurs in rice-growing countries worldwide and has recently been reported in Italy and Africa as well as in other Asian countries (Niehaus et al., 2017). It is a seed-borne fungal disease that can be controlled with the use of seed disinfectants; however, the incidence of the disease is increasing due to the recent introduction of sustainable eco-friendly farming practices and increase in rice transplanting. To control bakanae, the primary measures include the use of clean and healthy seeds, cultivation of resistant varieties, and seed disinfection with fungicides. However, frequent genetic mutations occur and result in the emergence of pathogenic variants in the field. Even if there is a resistant rice variety, such resistance can be broken down due to the high variability and fast evolving fungal populations. In addition, emergence of pathogens resistant to seed disinfectant fungicides has posed as an obstacle to effective control (Kang et al, 2017; Shin et al., 2008a).
Therefore, this review aims to present a strategic direction to responsive measures to bakanae by understanding the epidemiological aspects of the disease and evaluating the impact of climate change on the disease incidence in Korea. To this end, existing research on the pathogen, host plant, and environment—elements affecting the pathogenicity of bakanae—was summarized. In particular, various methods to control bakanae have been described and categorized into chemical, biological, cultural, resistant variety, and physical methods. Finally, we present various systematic applications for the integrated management of bakanae, as it is predicted to remain a major disease under climate change.
Bakanae in Korea
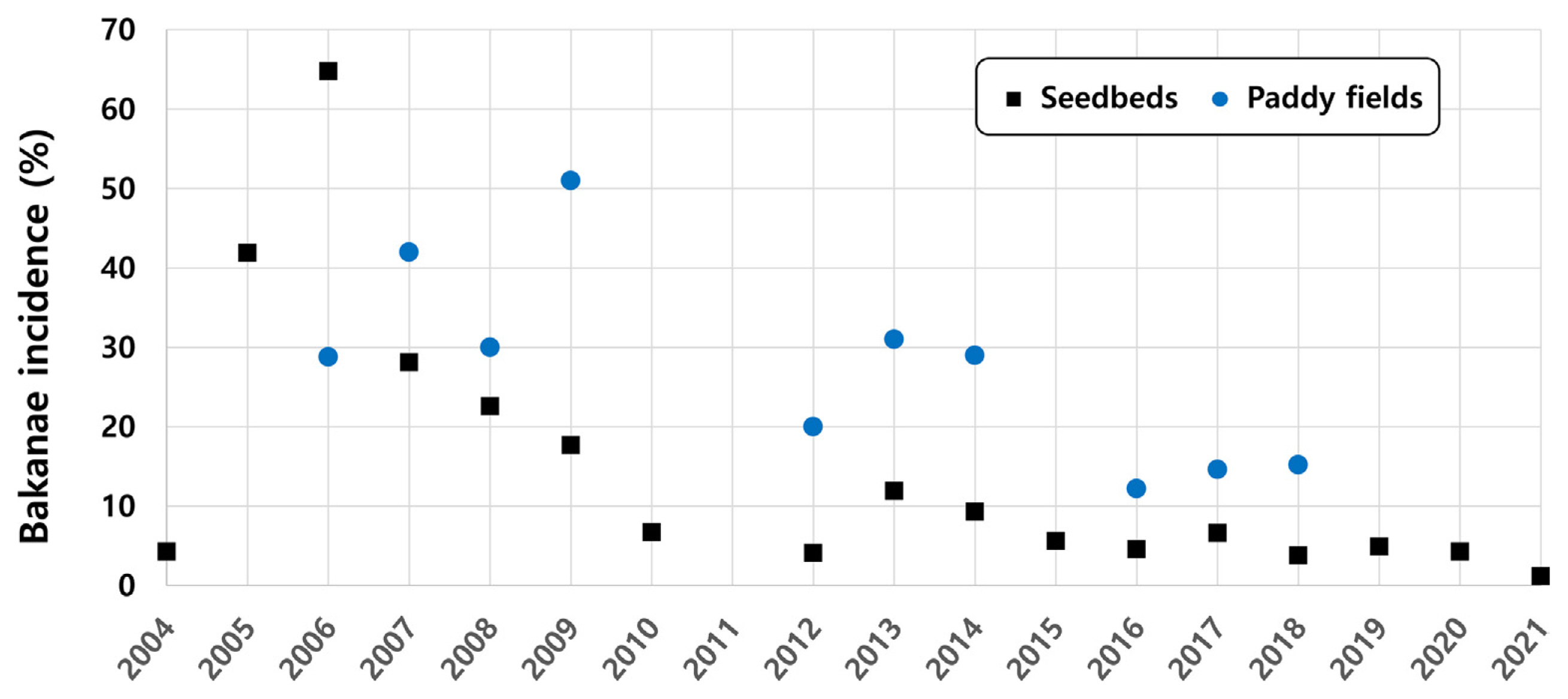
In Korea, bakanae was not a problematic disease in the 1960s because of the prevalent use of organic mercury pesticides. Even after 1978, when the use of organic mercury pesticides was banned, the occurrence of bakanae did not cause significant problems on farms because systemic seed disinfectants such as prochloraz were introduced (Park et al., 2003). However, the incidence of bakanae gradually increased since 2003. In 2006, a large outbreak of bakanae occurred, with its incidence in the seedbeds and paddy fields reaching 64.8% and 28.8%, respectively (Han, 2007; Rural Development Administration, 2007) (Fig. 1). Such outbreaks of bakanae were due to changes in farming practices (e.g., early farming and box seedling) and the emergence of fungicide-resistant pathogens (Lee et al., 2010; Shin et al., 2008b). After the introduction of seed disinfectant fungicides and the hot water immersion method, the disease incidence gradually decreased. However, in 2013, the incidence of bakanae in the seedbeds and paddy fields increased to 11.9% and 31%, respectively (Fig. 1). Given that the causal agent of bakanae, Fusarium fujikuroi, favors warm and humid climates (Manandhar, 1999; Matić et al., 2021), the variable incidences since 2012 were likely to be affected by weather variabilities during the cropping seasons. Moreover, from 2022, the Korea Seed & Variety Service distributes only rice seeds that are not disinfected but promotes the hot water immersion method. Although seed disinfection using the hot water immersion method or fungicides is widely performed by farmers, there will be a higher risk of rice bakanae if farmers fail to follow the recommended disinfection process.
Epidemiology of Bakanae
Disease cycle
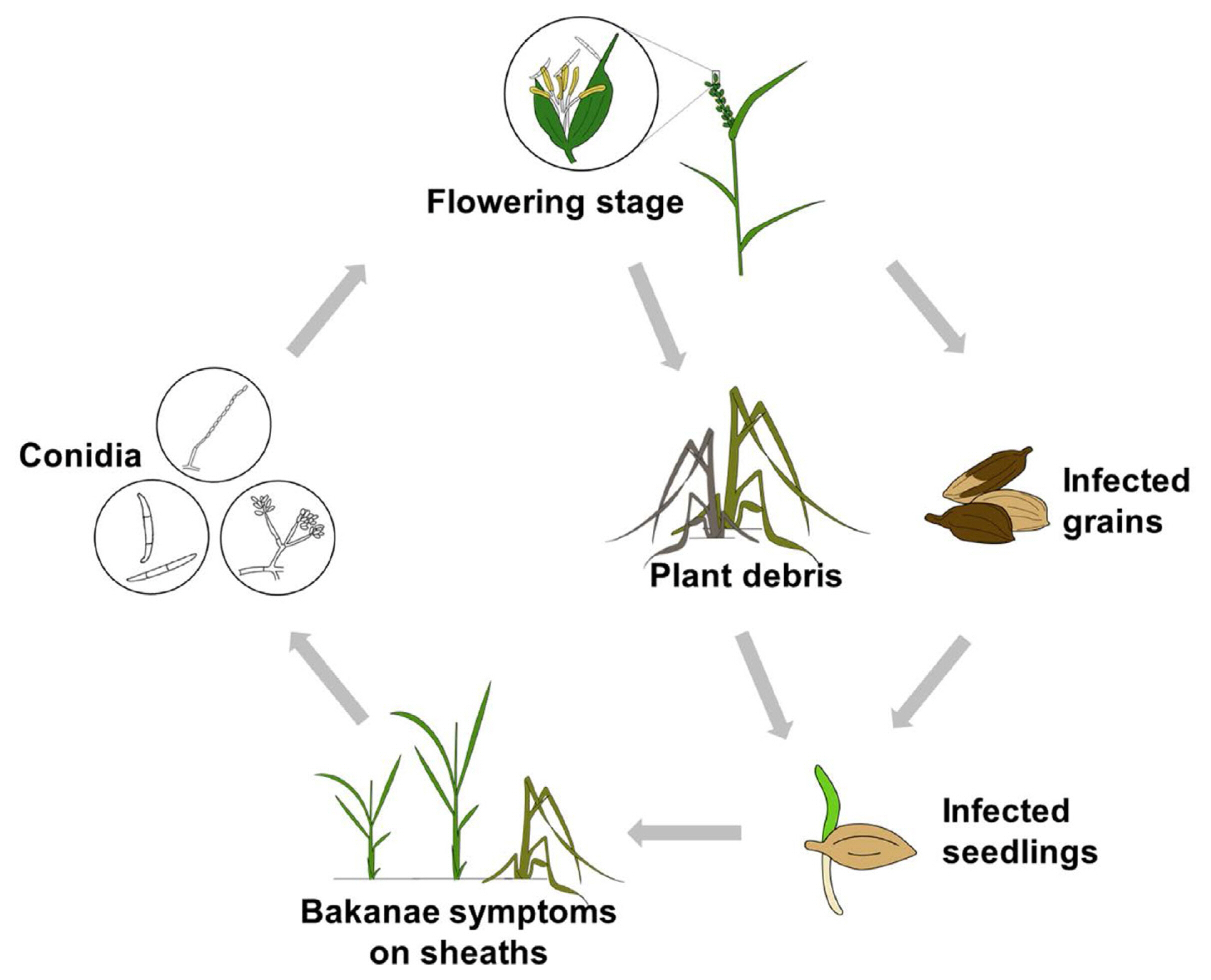
Bakanae, caused by F. fujikuroi, is a monocyclic disease (Fig. 2). It is generally known that bakanae is transmitted through seeds that are infected from the previous cropping season (Desjardins et al., 1997). The bakanae pathogen may also survive and be present as inoculum in overwintering plant debris or in soil (Kanjanasoon, 1965; Su et al., 1979; Sunder, 1997). In addition, Choi et al. (2011) isolated and confirmed the pathogenicity of the bakanae pathogen from barnyard grass, suggesting the possibility of barnyard grass as an alternative inoculum source.
When the infected seeds are sown in the next season, the infected seedlings are at least 1.5 times taller, with the appearance of leaf yellowing, than non-infected ones in rice nurseries. These plants wither after transplanting (Eğerci et al., 2021). Although no symptoms are shown at the time of transplanting, the infected plants either wither before the heading stage or remain standing with stems and sheaths covered with large numbers of white or pink conidia (Lee et al., 2011). Conidia formed at the tillering stage are dispersed by rain splash or wind during the flowering stage and infect floral organs, infecting seeds at later stages (Ou, 1985). Conidia can invade the inner and outer surfaces of rice glume, embryo, or endosperm. When the seeds, whose embryos or endosperm are infected, are sown in the following year, most seedlings wither while germinating (Manandhar, 1999).
Typical symptoms in the rice paddies include elongated growth, yellowing of leaves, stunting, seedling rot, and barren ear (Fig. 3). Nur Ain Izzati Mohd et al. (2008) reported that the 72-h period after pathogen inoculation is the most important period for successful infection of F. fujikuroi. The bakanae symptoms generally appear between 5 and 40 days post-inoculation (dpi). Symptoms such as yellowing and stunting are observed at 5-10 dpi. In the case of severe infection, the plant withers between 10 and 40 dpi. Occasionally, infected seedlings show late disease development in paddy fields. When they reach the panicle emergence stage, empty panicles are often formed (Chan et al., 2004).
Pathogen
F. fujikuroi is a plant pathogenic fungus belonging to the genus Fusarium of Ascomycota. The fungus is widely distributed worldwide in rice-growing countries, and is differentiated in terms of taxonomy and morphology according to the regions of distribution. F. fujikuroi is a member of the F. fujikuroi species complex (FFSC), which includes at least 50 phylogenetic species and is grouped into three clades: the American, African, and Asian clades (Niehaus et al., 2016). Three members of the FFSC, F. fujikuroi, F. proliferatum, and F. verticillioides, are reported to cause bakanae of rice. Among them, F. fujikuroi is found only in Asia (Wulff et al., 2010).
F. fujikuroi causes bakanae symptoms as a result of high gibberellic acid (GA) production in host plants (Rodrigues et al., 2012). Even in minimal amounts, GA has various physiological effects on plants, such as breaking dormancy and promoting germination in seeds, and fruit maturation. F. fujikuroi produces GA3, which functions as a general plant growth hormone, which may disrupt the hormonal regulation of growth and development in infected plants. However, GA production is not essential for fungal growth and development. Gene-disrupted mutants deficient in GA biosynthesis showed the colonization of pathogens in the plant cells and were able to infect rice roots (Wiemann et al., 2013). This is because F. fujikuroi produces a wide variety of secondary metabolites as well as GA, many of which are harmful to the host plant. Representative metabolites include mycotoxins (fumonisin, fusarins, and fusaric acid) and pigments (bikaverin and fusarubins) (Janevska and Tudzynski, 2018). It was previously reported that fumonisin can cause physiological stress including browning of stalks, leaves, and ears of rice in the flowering stage (Kushiro et al., 2012).
In addition, a recent study reported that secondary metabolites of F. fujikuroi are involved in the differentiation of pathotypes. Niehaus et al. (2017) characterized the secondary metabolite profiles in eight different F. fujikuroi strains exhibiting different symptoms (elongation or stunting/withering). The strain B14 with stunting symptoms rarely produced GA but produced high levels of fumonisin and had additional polyketide synthase genes compared to other isolates. Choi et al. (2018) investigated the distribution of FFSC in major food crops of Korea, such as rice, barley, and maize, and found that F. fujikuroi is dominant in rice samples with a distribution of approximately 50%. All identified F. fujikuroi isolates expressed FUM1, the gene encoding a polyketide synthase essential for the biosynthesis of fumonisin, and 76% of the isolates produced high levels of fumonisin, indicating that F. fujikuroi distributed in Korea may be able to cause stunting in addition to elongation.
Environment
Seedling stage
In the seedling stage, soil moisture is the most important environmental factor for successful pathogenesis. Latif et al. (2021) investigated the correlation between soil moisture conditions and bakanae incidence in seedbeds, and found that the incidence of bakanae decreased by approximately 10 times in wet seedbeds compared to dry seedbeds. In another study, the severity of bakanae also decreased with increasing soil moisture, and 5 times of severity reduction was observed under 100% soil moisture conditions compared to 30% soil moisture (Yadav et al., 2020). This may indicate that anaerobic conditions caused by excessive moisture are unfavorable to F. fujikuroi. Temperature is another important factor for bakanae development in the seedling stage. Strong positive correlations between disease incidence and soaking temperature (r = 0.8757) and disease incidence and sprouting temperature (r = 0.9570) were observed at 28-34°C (Chan et al., 2004). Coincidentally, the optimum temperature range for rice germination is 20-35°C. This could mean that the dry environment in seedbeds and the optimum temperatures at soaking and sprouting are important factors for the symptom development of bakanae.
Tillering and flowering stage
In these stages, temperature seems to be the most important factor for bakanae infection. In general, the optimum temperature for the growth of F. fujikuroi is 27-30°C, while a high temperature of 35°C is considered the most optimal temperature for bakanae infection (Bashyal, 2018). Furthermore, the disease symptoms are hardly observed at lower temperatures. For example, Bashyal et al. (2014) reported that the characteristic symptoms of bakanae are not manifested at 20°C, although the pathogen is isolated from the plant. Temperature is also an important determinant in the development of bakanae symptoms. Piombo et al. (2020) reported that the development of symptoms, even for the same strains, differed depending on temperature. Symptoms of elongation were observed at 31°C, but not at 22°C. When rice was inoculated at 31°C, the expression of the GA gene cluster significantly increased in the pathogens. Recently, the combined effect of CO2 with temperature on the incidence of bakanae was evaluated. Matić et al. (2021) reported that the disease index and death incidence of rice inoculated with the pathogen were elevated under high temperature and CO2 concentration, with solid support from genetic analysis showing the specific conditions favorable for disease progression. In the disease cycle of bakanae (Fig. 2), the most important event is the initial floral infection by dispersed conidia of the pathogen in the flowering stage. This is because the infected seeds at the flowering stage serve as the primary inoculum for the following season. At this stage, the most critical environmental factors are temperature and humidity, which affect fungal infection during sporulation, germination, and colonization of floral parts by the pathogen; and wind and rain, which aid the dispersal of conidia. The grain infection is known to increase at high temperature and insufficient insolation during flowering stage (Singh et al., 2019; Hino and Furata, 1968; Takeuchi, 1972). The aerial diffusion of conidia increased under heavy rain and strong wind conditions (Matić et al., 2017). Furthermore, the rate of infected seeds increased notably in the wet season with heavy rainfall (Manandhar, 1999).
Post-harvest stage
After the rice harvest, pathogens overwinter until the following season. Pathogens can survive up to 280 days in dry soil with a moisture holding capacity (MHC) of 10% (Su et al., 1979). However, as the MHC increases, the length of survival for the pathogen is reduced to 115 days under 45% MHC and 45-70 days under 100% MHC conditions.
Potential Epidemics under Climate Change
Modeling efforts
Based on the epidemiological understanding of bakanae, risk factors affecting the incidence of rice bakanae can be selected, and modeling and simulation of the disease incidence can be performed based on the interaction of multiple risk factors. However, a thorough literature search showed that a predictive model for the simulation of grain infection caused by bakanae during the rice cultivation period had not been studied. Instead, Lee (2009) developed a regression analysis model to estimate rice yield reduction using the level of bakanae incidence at different growth stages of two rice varieties in paddy fields in Korea. Chung et al. (2016) detected rice seedlings infected with bakanae using a support vector machine (SVM) model for the classification of infected seedlings. Because the SVM model allows objective and efficient classification of infected seedlings, it can be utilized for monitoring bakanae incidence and the development of bakanae-resistant rice varieties.
Based on the comprehensive review of previous studies, weather conditions during the flowering stage are expected to have a significant effect on rice bakanae infection (Bashyal, 2018; Bashyal et al., 2014; Manandhar, 1999; Matić et al., 2017, 2021; Piombo et al., 2020; Singh et al., 2019). Considering that rice grains infected during the rice flowering stage determine the initial incidence level of bakanae in the following year, a prediction model can be developed for simulating bakanae infection based on meteorological information during the flowering stage.
The EPIRICE simulates various diseases in rice (rice blast, sheath blight, bacterial blight, brown spot, tungro disease, etc.) through a simple and generalized algorithm of infection (Savary et al., 2012). The infection algorithms used in the EPIRICE can be found in the reports of Savary et al. (2012) and Kim et al. (2015). A simple bakanae infection model was developed based on the infection algorithm of EPIRICE, considering the environmental conditions and timing of grain infection during the rice cultivation period. The output of the model represents the ratio of infected grains to uninfected grains in the flowering stage. In the bakanae infection model, grain infection by airborne conidia occurs intensively for 20 days during the flowering stage. Two in vitro experiments under different temperature conditions revealed a simplified rule of the bakanae intensity-temperature relation as 69% at 25°C, 73% at 30°C, and 28% at 35°C, which was used as a temperature-response function in the bakanae infection model. The resulting infection model was subsequently calibrated based on three independent bakanae incidence data collected from the Rural Development Administration plots in 2021 and its applicability was ensured. The development of the bakanae infection model will be published in the near future. In this review, the impact assessment of climate change on rice bakanae in the Korean Peninsula was conducted using the bakanae infection model.
Climate change impact assessment
According to the disease cycle of rice bakanae (Fig. 2), seeds that were infected from the previous cropping season transmit the disease at the following season, indicating that the grain infection taking place at the flowering or maturing stage, and further the climatic conditions affecting the infection during that stage, is very critical (Desjardins et al., 1997). Because bakanae is a rice disease favoring high temperature, humidity, and CO2 concentration, there is a high possibility that it increases with the warm and humid conditions prevailing under climate change (Manandhar, 1999; Matić et al., 2021; Piombo et al., 2020). According to Bashyal (2018), the optimal temperatures for the infection and development of bakanae is 27-30°C and 35°C, respectively, which are warmer than the optimal temperatures (approximately 29 °C) for the development of rice bacterial leaf blight, a well-known thermophilic disease (Horino et al., 1982). Therefore, it is important to understand the projected behavior of bakanae under climate change using the infection model and climate change scenarios from global climate models (GCMs).
A climate change impact assessment to project the potential future incidence of bakanae was conducted by combining the bakanae infection model and climate change scenarios of the Coupled Model Intercomparison Project 6 (CMIP6). For this purpose, downscaled data of the SSP5-8.5 climate change scenarios of the CMIP6 models was used as input for the infection model. The SSP5-8.5 corresponds to a nominal anthropogenic forcing of 8.5 W/m2 by 2100, which reflects the continuously increasing trend of greenhouse gas emission in the face of limited global/national policy intervention (Olivier et al., 2017).
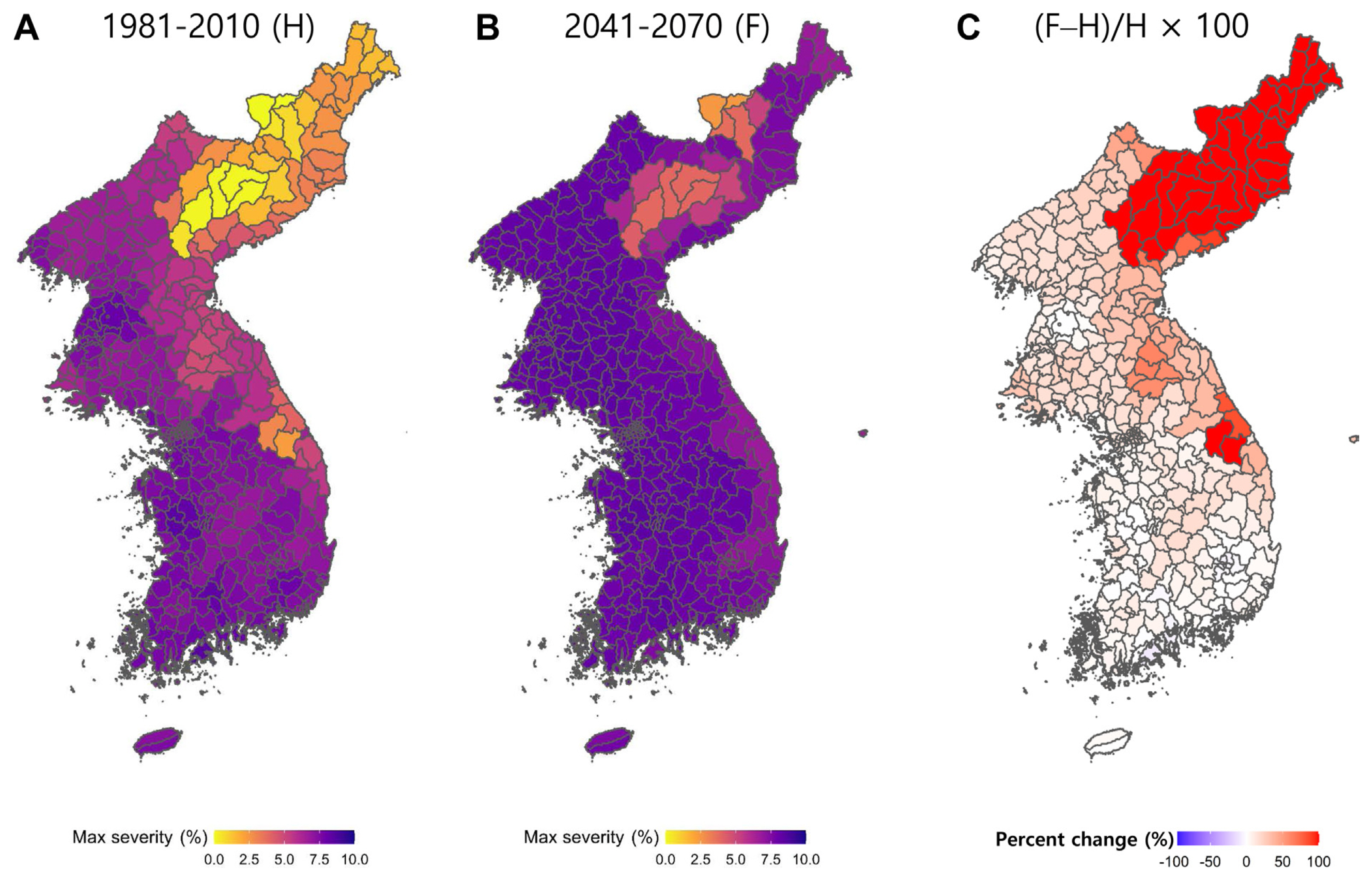
Briefly, bias correction and downscaling were conducted for the scenario data from eight GCMs (CanESM5, CNRM-CM6-1, GFDL-ESM4, INM-CM5-0, IPSL-CM6A-LR, MIROC6, MRI-ESM2-0, and UKESM1-0-LL) using simple quantile mapping (SQM), one of the most commonly-used bias correction methods (Cannon et al., 2015). Climate observation data from 87 Automatic Synoptic Observation System (ASOS) stations across the Korean Peninsula were obtained from the Korea Meteorological Administration (KMA) and used for SQM. Historical climate data for 1981-2010 and future climate data for 2041-2070 were used as inputs for the bakanae infection model, and changes in the ratio of infected grains, or the maximum severity of rice bakanae were simulated using the scenarios. The 30-year average values of the simulated ratio of infected grains were calculated for the respective historical and future periods, and these were then visualized on maps by spatially interpolating the point values over 415 administrative districts of the Korean Peninsula using the kriging method (Nelson et al., 1999), as shown in Fig. 4.
Simulations for the historical period (1981-2010) indicated that the climatic environment of the Korean Peninsula is conducive to rice bakanae in most regions, especially showing more intense epidemics in most provinces of South Korea and western provinces of North Korea. Among all administrative boundaries, Goheung-gun of Jeollanam-do in South Korea showed the highest ratio of infected grains at 8.449%. Also, Jeollanam-do, Chungcheongnam-do, Jeollabuk-do, Pyongyang, and Gyeongsangnam-do showed relatively high severities of bakanae. In contrast, Taehongdan-gun and Ryanggang-do in North Korea showed the lowest severity of 0.005%, and Ryanggang-do, Hamgyongnam-do, Pyongannam-do, and Hamgyongbuk-do, the northeast regions of North Korea, also showed relatively low severities. The simulation results for the future (2041-2070) showed that the maximum severity of bakanae increased in most regions of the Korean Peninsula compared to that in the historical period. Particularly, the northeast region in North Korea showed a significant increase in the maximum severity.
Based on the difference between the simulated severities of the historical and future periods, the percentage change in the risk of bakanae intensity in the future compared to that of the historical period was calculated. In all regions except for 18 administrative districts in North and South Korea, the bakanae severities were predicted to increase in the future. Taehongdan-gun in Ryanggang-do, North Korea, showed the largest increase among 39 administrative districts, where the changes between the historical and future periods exceeded 100%. In contrast, the administrative districts of Jeollanam-do, Gyeongsangnam-do, Daegu Metropolitan City, and Gyeongsangbuk-do showed relative decreases in the maximum severity of bakanae, and the district with the greatest decrease was Goheung-gun in Jeollanam-do, which showed a reduction of approximately 11.02%.
Overall, as previously hypothesized based on literature review, our study showed that rice bakanae severity is projected to increase in the future under climate change, showing an increasing trend overall. The percent change was particularly larger in the northeast region of North Korea. These results re-emphasize that the global warming-induced increase in the risk of bakanae in the Korean Peninsula should encourage scientists and policy makers to prepare for integrated bakanae management using the effective combinations of multiple disease management options ranging from chemical control to biological, cultural, resistant variety, and physical control measures.
Management of Bakanae Disease
Chemical control (fungicide resistance)
Chemical fungicides for controlling bakanae can be categorized, according to their mode of action, into benzimidazoles, demethylation inhibitors (DMIs), succinate dehydrogenase inhibitors (SDHIs), and strobilurin fungicides.
Benzimidazole fungicides include benomyl, carbendazim, thiabendazole, and thiophanate-methyl; these fungicides are involved in β-tubulin biosynthesis (Li et al., 2021). Carbendazim (Bavistin 50WP) of benzimidazole fungicides and dithiocarbamate (Greenzeb) of mancozeb fungicides have been reported to be the most effective (Hossain et al., 2015). The long-term application of the benzimidazole since the 1970s has led to the emergence of fungicide-resistant strains in fields, resulting in lower control efficiency against the bakanae pathogen. In Japan, strains resistant to thiophanate-methyl and benomyl emerged in the 1980s (Ogawa and Takeda, 1990), and strains resistant to carbendazim bearing mutations in the β2-tubulin gene were reported in China (Chen et al., 2014).
DMIs target cytochrome P450 and inhibit the metabolism of fungi, including the ergosterol biosynthesis pathway. Representative DMIs include difenoconazole, ipconazole, pefurazoate, prochloraz, penconazole, tebuconazole, propiconazole, and triflumizole. Prochloraz is an imidazole-class sterol demethylation inhibitor and has been used as a seed disinfectant for a long time in Korea. However, in 2010, F. fujikuroi isolates resistant to prochloraz and benomyl + thiram fungicides were reported (Kim et al., 2010). In addition, Lee et al. (2010) reported strains resistant to prochloraz + tebuconazole, prochloraz + benomyl, and tebuconazole + benomyl.
SDHI fungicides can inhibit ATP production by blocking the electron transport of the fungal mitochondria (Sierotzki and Scalliet, 2013). Representative active ingredients of SDHI fungicides include pyrazole-4-carboxamide, carboxin, and thifluzamide. Strobilurin-class fungicides for the control of bakanae include azoxystrobin, kresoxim-methyl, and orysastrobin (Nofiani et al., 2018). In China, strains resistant to phenamacril, a fungicide of cyanoacrylate compound, emerged within 5 years of application of the fungicide (Hou et al., 2018; Li et al., 2008). A recent study reported that resistance of pathogens to phenamacril increased from 18% in 2017 to 47% in 2018 (Wu et al., 2019).
The frequent emergence of fungicide-resistant strains is because genetic crosses between F. fujikuroi strains are relatively easy (Dyer et al., 2000). To tackle fungicide resistance, combined application of more than one chemical fungicide with different mechanisms of action has been proposed in recent reports (Gisi et al., 1985). For example, in a greenhouse experiment, it was reported that treatment with a 2:1 mixture of phenamacril and ipconazole could control almost 100% of bakanae (Li et al., 2017). Park et al. (2009) reported that mixed use of prochloraz and fludioxonil increased the efficiency of seed disinfection and showed an excellent control effect at 96.7%. Strains resistant to benomyl or thiram are sensitive to DMI fungicides such as triflumizole, propiconazole, ipconazole, and prochloraz. Therefore, combinations of these fungicides are reported in many studies for antifungal cocktail treatment (Bagga et al., 2007; Ghazanfar et al., 2009; Karov et al., 2009; Ora et al., 2015; Sharma, 2006; Tateishi et al., 1998; Waller, 1987).
In Korea, various fungicides are commercially available for rice bakanae (listed in Table 1). Total 38 active ingredients (combination or alone) are registered for seed and seedling treatment based on the pesticide safety information system (Rural Development Administration, 2022). In general, seed/seedling treatment can be divided into (1) seed dressing, (2) seed soaking, and (3) seedling box treatment. Seed dressing, is conducted by applying chemicals to the seeds. In the past, effective combinations for seed dressing include prochloraz + carbendazim, mancozeb + benomyl, and carboxin + thiram + iprodione (Garibaldi, 1985). However, fungicides including fludioxonil, carboxin, or pyribencarb are used for seed dressing in present Korea. Carbendazim was reported to be effective for application in the seed soaking treatment method (Aurangzeb et al., 1998), but fungicides including carbendazim for seed soaking treatment are no longer commercially used in Korea. The active ingredients used in Korea for fungicides of seed soaking treatment include prochloraz + triflumizole and pyribencarb + triflumizole. A method frequently used along with the seed treatment method is seedling box treatment with bavistin and benomyl in other countries (Sharma, 2006). In Korea, 22 out of 37 active ingredient combination (or alone) are registered for seedling box treatment (Rural Development Administration, 2022), and these fungicide combinations includes probenazole + imidacloprid, azoxystrobin + clothianidin, and isotianil + penflufen + clothianidin.
For seed dressing, combining other types of control methods with chemical control is also an effective technique. As a representative combination, heat treatment, a physical control method, can be combined with chemical control. The control effect is reportedly enhanced when heat treatment is performed along with chemical treatment of carbendazim, mancozeb, iprodione + propiconazole, or carboxin + thiram. Titone et al. (2003) reported that heat treatment at 72°C for 5 min combined with chemical treatment of carbendazim showed a control effect of 98%. In a recent study on sustainable and eco-friendly pest control, the use of slightly acidic hypochlorous acid water exerted a potent antimicrobial effect to the pathogen, resulting in effective seed disinfection and disease control performance (Goo and Koo, 2020). In addition, chitosan oligosaccharides and ethylenediaminetetraacetic acid showed excellent fungicidal effects (Kim et al., 2016).
Various control methods can be applied for parts of the rice plant in other growth stages as well as seeds and seedlings. It has been reported that the use of benzimidazole as a fungicide for foliage treatment inhibited mycelial growth and reduced the incidence of bakanae (Biswas and Das, 2002). In addition, treatment with benomyl, carbendazim, and propiconazole applied across the plant was reported to effectively prevent infection and disease transmission (Hajra et al., 1994; Sasaki, 1987). The fungicide combinations for foliage treatment used in Korea include hexaconazole + tiadinil, ferimzone + hexaconazole, and fenoxanil + thifluzamide, which were recently registered in 2019-2021 (Rural Development Administration, 2022).
Biological control
Prokaryotes
One of the common methods for biological control of bakanae is applying various microorganisms isolated from the rhizosphere or soil of the target crop. In 1986, Rosales et al. (1986, 1993), for the first time, isolated bacteria belonging to the genera Bacillus, Pseudomonas, and Serratia from a healthy host plant and its surrounding rhizosphere using an in vitro method and confirmed that the isolates had a control effect on bakanae. Among these microorganisms, effective strains were selected for seed treatment and field trials for three years, and the result confirmed the control effect in field applications (Rosales and Mew, 1997). Luo et al. (2005) also isolated effective gram-positive bacteria against bakanae in China, such as Bacillus megaterium, B. subtilis, B. pumilus, B. cereus, B. licheniformis, and Brevibacillus brevis. In addition, Khaskheli et al. (2020) confirmed that endophytic bacteria such as those belonging to genera Bacillus, Fictibacillus, Lysinibacillus, Paenibacillus, Cupriavidus, and Microbacterium isolated from rice roots have biocontrol activity against F. fujikuroi. However, the underlying effects of the active ingredients were not analyzed.
As microbiological studies were combined with biochemistry, molecules with antifungal activity were identified. Nagaraj Kumar et al. (2007) extracted chitinase, β-1-3-glucanase, siderophore, salicylic acid, and hydrogen cyanide from Pseudomonas fluorescens and B. subtilis. These chemicals which can degrade mycelia of F. fujikuroi are effective in controlling bakanae. Zhang et al. (2012) extracted an antifungal lipopeptide called iturin A, which leads to abnormal mycelia in F. fujikuroi, from Bacillus amyloliquefaciens. Surfactin A obtained from Bacillus strains also showed the biocontrol activities against the bakanae pathogen (Sarwar et al., 2018). Apart from enzymes and non-volatile organic compounds (VOCs), VOCs have also been identified as effective molecules to control the bakanae pathogen. The 2-methylisoborneol produced by Streptomyces alboflavus was confirmed to inhibit the growth of F. fujikuroi (Wang et al., 2013).
Some bacteria show indirect antifungal activities by enhancing immune systems of rice. For example, in Korea, Bacillus oryzicola, an endophytic bacterium isolated from paddy rice roots, showed a significant effect in controlling bakanae when it was used to drench the rice rhizosphere. Via a bioassay of detached rice leaves, B. oryzicola was confirmed to suppress bakanae through induced systemic resistance (Chung et al., 2015; Hossain et al., 2016). Furthermore, Bacillus species in the rhizosphere and endosphere of rice elicited peroxidase activity in the crop, thereby increasing resistance to F. fujikuroi (Nawaz et al., 2022).
Eukaryotes
Among fungi, the genus Trichoderma is a well-known biocontrol agent producing enzymes (Ajijolakewu et al., 2017). Watanabe et al. (2007) reported that T. asperellum could control bakanae by degrading the cell walls of F. fujikuroi. In addition, compounds produced by Trichoderma showed antagonistic activity against F. fujikuroi, and T. harzianum was able to inhibit the growth of F. fujikuroi in in vitro conditions (Bhramaramba and Nagamani, 2013; Halim et al., 2015). High enzyme production in Trichoderma spp. (amylase, cellulose, lipase, pectinase, and protease) suggested that Trichoderma spp. exhibit a significant inhibitory effect on the growth of F. fujikuroi compared to other soil fungi (Rajathi et al., 2020).
Talaromyces spp. isolated from rice seedlings were also reported to be effective in the control of F. fujikuroi as a fungal symbiont (Kato et al., 2012). Fungal isolates belonging to the genus Phoma from the zoysiagrass rhizosphere were able to control bakanae and increase the fresh and dry weight of rice (Elsharkawy et al., 2014). Antagonistic activity similar to that of bacterial biocontrol agents was also found in the fungi. Khunnamwong et al. (2020) showed that Wickerhamomyces anomalus, a yeast strain isolated from the leaf tissue of rice, corn, and sugarcane, produces VOCs, fungal cell wall degrading enzymes such as β-1,3-glucanase and chitinase, and siderophores to exhibit antagonistic activity against F. fujikuroi.
Natural products
Natural products include plant essential oils, which contain terpenoid and phenolic compounds. These plant products exhibit antimicrobial activity. In Europe, they are widely applied as an effective alternative to synthetic antifungal agents for the control of seed-borne F. fujikuroi. In fact, treatment with thymol and eugenol over two years could effectively control F. fujikuroi (Mongiano et al., 2021).
Combination with other control methods
In the case of bacteria, Kazempour and Elahinia (2007) reported that simultaneous treatment of seeds with Rovral TS, a chemical fungicide, and antagonistic bacteria has a more significant control effect than individual treatment. On the contrary, among fungi, the treatment with Trichoderma alone exhibited a greater control effect than that of the combinations with conventional chemical pesticides (Pal et al., 2019). These conflicting results explained different metabolic mechanisms and environmental ecology between bacteria and fungi. In addition, Matić et al. (2014) reported that when Metschnikowia pulcherrima, Pichia guilliermondii, and Sporidiobolus pararoseus, which are antagonists to F. fujikuroi, were inoculated in seeds that were heat-treated at 60°C, the control effect was higher than in seeds that were individually treated with the biocontrol agents or heat.
Cultural control
Although bakanae is primarily seed-borne, it can also be transmitted through the overwintering inoculum in plant debris and in soil. However, it is reported that the inoculum in soil is rapidly reduced with decomposition of the host debris (Kanjanasoon, 1965; Sunder, 1997), and a high soil moisture content is known to reduce the bakanae incidence (Stodola, 1958). These findings may indicate that the soil is not the main route of infection in Korea, as the moisture content of the soil is maintained at a high level during the early growing season of rice. In the case of severe outbreaks of bakanae, stubble burning and removal of diseased plant debris as well as rotation with non-host plants may help reduce incidence by reducing the primary inoculum (Burgess et al., 1996; Gupta et al., 2004).
Because bakanae is mainly transmitted by infected seeds, the use of healthy seeds is the most useful and realistic method for prevention of the disease. Seeds should be harvested from uninfected paddy fields, and the method of selecting healthy seeds using salt water for sowing also helps in reducing the incidence of bakanae (Cother and Lanoiselet, 2002). A similar effect can be obtained by filtering out light seeds using an awn remover. In the seedling stage, the incidence of bakanae was drastically reduced when a paddy nursery was uprooted in standing water due to a reduced rate of root injury (Sunder et al., 2014). In addition, if seedlings showing typical symptoms of bakanae are identified during the seedling phase, they should be removed, thereby minimizing seed-borne infection.
The use of chemical fertilizers or compost may also have a direct or indirect effect on the incidence of bakanae. The most recent study reported a low incidence when no element was used excessively or insufficiently in the applied fertilizer (Kumar et al., 2022; Mandal and Chaudhuri, 1988; Pavgi and Singh, 1964). When the disease incidence was examined by applying various compositions of N, P, K, Zn, and Fe, the incidence was lower when these elements were not applied in excess (Kumar et al., 2022). In addition, the incidence was lower when not only NPK but also trace elements such as Zn and Fe were applied at the recommended concentrations. Therefore, the incidence of bakanae can be reduced by optimizing the amount of fertilizer for each field by conducting a soil analysis before starting rice cultivation every year.
Since bakanae occurs frequently at high temperatures of ~35°C, lowering the temperature during the outbreak period by delaying the cultivation period of the crop will also contribute to reducing the disease incidence (Bagga et al., 2007; Bal and Biswas, 2018; Heaton and Morschel, 1965). In India, it was confirmed that the earlier the rice was transplanted, the higher the disease incidence. This is because when rice is transplanted earlier, the temperature and relative humidity are more suitable for fungal growth (Sandhu and Dhaliwal, 2016). Therefore, geographical characteristics should also be considered to set optimum, local-specific management programs based on cultural control measures.
Resistant varieties
Molecular mechanisms of rice bakanae resistance
Research based on transcriptome analysis has been performed to reveal the mechanism of host resistance to bakanae. Ji et al. (2016) performed a comparative transcriptome analysis using resistant (93-11) and susceptible rice varieties (Nipponbare); in the resistant variety, an increase in the expression of defense-related genes WRKYs and MARK was confirmed. The Kyoto Encyclopedia of Genes and Genomes identification of Selenio, a bakanae-resistant variety, confirmed that genes such as Ca2+-dependent protein kinase, JASMONATE ZIM-DOMAIN-like, CEBiP, CERK1, and MYC2 are involved in the resistance mechanism (Spadaro et al., 2018). These genes were enriched in gene ontology terms associated with chitin, jasmonic acid biosynthesis, and plant-type hypersensitive response. When GA production was inhibited, the Selenio cultivar activated the jasmonic acid metabolic pathway, likely exhibiting hypersensitive responses to pathogens. Moreover, the Selenio cultivar induces an increased production of rice phytoalexins, whereas Dorella cultivar, a susceptible variety, induced the production of GA and abscisic acid, but inhibited jasmonic acid biosynthesis, thereby producing only a low level of phytoalexins (Siciliano et al., 2015). These findings indicate that the host plant becomes resistant to bakanae through the systematic up-regulation of resistance genes such as WRKY transcription factor, MAPK cascades, and some cytochrome P450s.
Bakanae resistant varieties in Korea
Typical methods for controlling rice bakanae include treatment of chemical fungicide; however, the increasing emergence of fungicide-resistant isolates has been reported (Lee et al., 2010). Thus, the cultivation of a disease resistant variety is a fundamental solution to bakanae, which can enhance the cultivation stability and reduce costs from other control measures. Sang et al. (2014) screened 33 japonica varieties for bakanae resistance in Korea. Nampyeong and Saenuri were identified as resistant varieties, and nine varieties including Daecheong and Hopum were identified as susceptible varieties. In addition, as a result of disease resistance screening using 254 rice germplasm including domestic and foreign japonica, Tongil-type, and indica varieties, a fully resistant variety to bakanae was not identified. Although some of the varieties exhibited resistance similar to that of Nampyeong, 131 varieties were identified as susceptible varieties (Hur et al., 2016).
In addition, Lee (2022) screened major domestic varieties and foreign varieties for bakanae resistance. Among the 66 varieties screened, Anpyeong, Samgwang, Nampyeong, and Misochan showed normal growth with dark green color and strong resistance with no typical symptoms of bakanae. Youngjin, Saenuri, Aranghangchal, and Josaengheugchal showed moderate resistance presenting with a light green color and elongation. In addition, Inwol, Shingwang, Milyang313, and MY299BK were discovered or developed as varieties showing resistance. Ilmi, Jinkwang, Misojinmi, Sangju, and Daean were varieties whose growth was almost stopped or that showed complete withering. Among the varieties mainly cultivated in Korea, Shindongjin, Ilpum, Chucheong, Younghojin, Saeilmi, Dongjinchal, and Chindeul also showed little resistance to bakanae.
Examples of varieties resistant to bakanae that can be supplied in Korea include Anpyeong, Nampyeong, Samgwang, Misochan, Saenuri, Youngjin, Josaengheugchal, and Aranghangchal. In particular, Anpyeong, Nampyeong, Samgwang, and Misochan were reported to exhibit strong resistance to bakanae (Lee, 2022), and therefore, yield loss due to bakanae can be reduced through selective cultivation of these varieties. However, the information on rice varieties supplied nationwide in 2021 showed that most varieties except ‘Samgwang,’ the second most cultivated variety, showed little or no resistance to bakanae (Table 2). As a result, whenever an environment is conducive to the occurrence of rice bakanae, there is a high possibility of an outbreak.
Development of resistant varieties using the bakanae resistant quantitative trait locus
Cultivation of resistant varieties has limitations for various reasons (availability, rice quality, optimal environmental conditions, the quick emergence of new pathogenic strains, etc.) (Lee et al., 2021). Therefore, many different quantitative trait locus (QTL) related to bakanae resistance have recently been identified through QTL mapping, which can be used to perform marker-assisted selection in rice cultivation and resistance mechanisms to bakanae (Lee et al., 2019; Park, 2020). Most QTLs conferring bakanae resistance were identified on chromosome 1 and qBK1z, qBK1.2, and qBK1.3 are localized in similar regions of different resistant varieties (Lee et al., 2021). Yang et al. (2006) performed QTL analysis for bakanae resistance by artificial inoculation using a japonica/indica double haploid population derived from Chunjiang06/TN1; two QTLs were identified on chromosomes 1 and 10. Fiyaz et al. (2016) used a population derived from the indica rice parent Pusa 1342, and a total of three QTLs governing resistance to bakanae were identified (qBK1.1, qBK1.2, and qBK1.3) on chromosomes 1 and 3. In China, two resistance QTLs were reported on chromosomes 1 and 10 using the Chunjiang 06/TN1 doubled haploid population (Yang et al., 2006).
In Korea, QTL analysis were performed among domestic resistant cultivars and qFfR9, a major QTL, was discovered on chromosome 9 using the rice variety ‘Samgwang’ (Kang et al., 2019). In the case of ‘Shingwang’, qBK1, a resistant QTL, was found on chromosome 1, and an additional QTL, qBK1WD, was located from 13.54 to 15.13 Mb of chromosome 1 of ‘Wonseadaesoo,’ a resistant gene source of the japonica variety. A QTL from ‘Nampyeong’, a resistant variety, was reported at 87.9-91.7 cM of the same chromosome 1 (Hur et al., 2015; Lee et al., 2018). Based on the reported QTLs, ‘Anpyeong’ and ‘MY299BK’ were developed from a cross with ‘Shingwang,’ a variety with qBK1 (Jo et al., 2020).
Among domestic varieties with resistance to bakanae, ‘Guwangdo,’ ‘Wonsanchalbyeo,’ ‘Hongdo,’ ‘Gangwondo,’ ‘Akudichal,’ ‘Baekcheon,’ ‘Ssalbyoe,’ ‘Annamjo,’ ‘Parkssina,’ ‘Gangneungdo,’ and ‘Sukna’ showed greater resistance than Shingwang rice (Gwon and Jin, 2018). Therefore, if a diverse range of QTL other than the previously reported ones can be discovered from the domestic varieties, they could be utilized as useful sources for the development of bakanae resistant varieties in the future.
Physical control
The pathogen was destroyed when infected seeds were buried in snow or subjected to low temperatures of 5°C (Sasaki, 1987). Thermotherapy, in which the pathogen is destroyed by immersing the seeds in water at approximately 60°C for 10 min, was also reported to help prevent the occurrence of rice bakanae (Miyasaka et al., 2000; Yamashita et al., 2000). Matić et al. (2014) reported that the disease index showed a significant decrease to a similar level at 60, 65, and 70°C but the seed germination rate decreased as the temperature increased. Therefore, the authors suggested that 60°C is the appropriate temperature for effective thermotherapy. In Korea, the use of thermotherapy with hot water is mainly promoted for controlling bakanae. Recently, research was carried out on seed disinfection by irradiating seeds with atmospheric plasma (Ochi et al., 2017). When the infected seeds were sterilized using atmospheric plasma irradiation, the incidence and disease index of bakanae were decreased. The production of hydrogen peroxide in seeds during plasma irradiation is believed to be an underlying sterilization mechanism.
Conclusions and Way Forward
Projection using climate change scenarios indicated that the incidence of rice bakanae will increase or at least remain at the same level in the future (Fig. 4). To minimize the potential risk of bakanae disease in the future, it is very important to understand the epidemiological nature of target disease at first. Comprehensive considerations of various factors, such as the characteristics of the disease cycle, pathogen, and environment, are presented in this review. Based on the epidemiological understanding, a full inventory of management options, which are presently available or need to be developed soon, firstly in Korea and also in other countries, were identified. This review provided most available bakanae management options, encouraging the use of the most effective combinations of multiple options ranging from chemical control to biological, cultural, resistant, and physical control measures.
Control measures should be applied accordingly to the variety, timing, and economic threshold level in the context of cultivation site. Although treatment with chemical fungicides has shown some promising results to control bakanae, certain drawbacks and limitations exist such as emergence of resistant strains, pesticide residues, and environmental pollution, possibly leading to harmful consequences for both the plant and humans. Therefore, with the aim of minimizing the side effects by reducing dependence on chemical fungicides, integrated disease management (IDM) is the ultimate goal.
In IDM, fungicides are used in combination with other approaches only when necessary for more effective and long-term control. First, an optimal amount of fertilizer should be applied, with no specific element in excess or in deficiency, through soil analysis during the land preparation of rice paddy fields. Next, healthy seeds should be obtained from the Korea Seed & Variety Service, while avoiding the use of uncertified seeds harvested in the farmer’s paddy field last year. In addition, if there was an outbreak of bakanae in the field last year, cultivation of rice varieties that are resistant to bakanae should mainly be considered. Examples of bakanae resistant varieties that can be supplied in Korea include Anpyeong, Nampyeong, Samgwang, and Misochan (Lee, 2022). By cultivating these varieties, yield losses from bakanae can be reduced with less fungicide applications.
Control effectiveness against bakanae increases when heat treatment or seed selection using salt solution is applied in addition to fungicide treatment (Titone et al., 2003). In addition, the use of mixtures of two types of fungicides are recommended. Combining fungicides with different modes of action allows control of the disease with relatively a small amount of fungicide and is more effective for limiting the development of fungicide resistance. In Korea, recently registered fungicide combinations for rice bakanae (Rural Development Administration, 2022) include azoxystrobin + clothianidin (registered in 2021), dichlobentiazox + thifluzamide + clothianidin (2022), dichlobentiazox + fipronil (2022), and azoxystrobin + pydiflumetofen (2022).
In the seedling process after seed disinfection, the optimum amount of seeds must be applied, and environments conducive to bakanae disease, such as high temperature and humidity, should be avoided. Seedlings of elongated growth need to be immediately removed as soon as they are detected in the seedbed. After transplanting to paddy fields, if any plants showing bakanae symptoms are identified, it is necessary to remove them immediately and apply the registered fungicide by spraying at the time of emergence of panicles. During decision-making on spray-application of fungicides, it is necessary to apply the optimal amount at an appropriate timing by identifying whether the meteorological conditions during the flowering stage are conducive to bakanae infection using the disease prediction model. In recent studies, new technologies for applying RNA interference for silencing a specific gene of a target pathogen have been developed, demonstrating the potential for the control of diseases caused by several Fusarium spp. (Šečić and Kogel, 2021). Therefore, further investigation on the target genes of F. fujikuroi must be encouraged to be able to ensure a comprehensive IDM (Koch et al., 2016; Shanmugam et al., 2017).
In recent years, with the increasing global interest in precision agriculture, there has been an increase in research on non-destructive early detection of crop diseases using hyperspectral sensors (Hernández-Clemente et al., 2019). Indeed, previous studies have investigated different types of plant stresses, pathogens, and plant diseases as follows: anthocyanin content (Steyn et al., 2002), water stress conditions (Hernández-Clemente et al., 2012), Xylella fastidiosa (Zarco-Tejada et al., 2018), Verticillium wilt (Calderón et al., 2015), and almond red leaf blotch (López-López et al., 2016).
The phenotypes of bakanae differ depending on the severity of infection. Most severely infected seeds wither during the germination process, whereas moderately infected seeds show elongated growth in seedbeds. Considering these characteristics, due to the nature of hyperspectral remote sensing that analyzes the characteristics of the target through spectral reflectance, some challenges in determining the presence or absence of pathogens inside the thick seed coats are expected. However, the detection of bakanae in fields after transplanting may be possible. Considering the elongated overgrowth in the field, which is a typical symptom of bakanae (Fig. 3), it is expected that the disease can be detected through the hyperspectral and thermal images. In the field, symptoms of bakanae are elongated and thinner seedlings and yellowing of leaves. This can be split into two parts: (1) elongated growth and thinning and (2) yellowing of leaves. First, the leaves of elongated overgrowth show a difference in canopy structure. The difference is expected to be determined by the estimation of the Fraction of Absorbed Photosynthetically Active Radiation (FAPAR) of the target plant. In addition, yellowing of leaves is expected to be detected through the chlorophyll/carotenoid index (ratio of transmittance at 931 nm to that at 653 nm, termed chlorophyll content index).
However, there are still limitations in detecting the bakanae symptoms using the remote sensing image techniques in plants. First, pigment, structure, fluorescence, and temperature are all secondary symptoms. In addition, since the physiological indicators described above may change depending on factors such as the environment and rice variety, it is difficult to determine whether the symptoms are caused by bakanae or other plant stresses. Therefore, a more realistic solution for the early detection of bakanae in the fields would be utilizing a bakanae infection model to understand the epidemiological behavior of bakanae influenced by daily weather conditions over the rice growth stages. Development of seedling infection and subsequent consequences of typical seedling symptoms such as elongation, stunting, and withering can be simulated using the infection model. Seedling infection algorithms can be developed based on empirical relations determining the multiple interactions between weather, pathogen, and host plant, leading to infection and symptom development in rice seedlings. Floral infection resulting in seed infection can also be predicted during the flowering stage based on governing weather conditions and relevant infection algorithms that have been determined from multiple in vitro and field experiments. The bakanae infection model we used for the climate change impact assessment in the study can be a starting point for further development, which later can be used in the early detection of bakanae in the fields for effective management of bakanae by farmers.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






