


The genus Medicago contains 85 species, two of which are shrubby and the others are herbaceous, including 20 perennials and 63 annuals, which are generally autogamous (Small and Jomphe, 1989). Most annual Medicago species attract agronomic interest for dryland forage production and for the suppression or reduction of nitrogen fertilizer application. Interest in these species is closely linked to their ability to naturally reconstitute themselves from one year to another. This property can lead to a permanent installation adapted to the vagaries of the Mediterranean zones, thus reducing the phenomenon of soil erosion. In addition, many Medicago species and subspecies also exhibit agronomic traits that would be desirable to introduce into cultivated alfalfa, such as pasture tolerance (rooting capacity and re-growth), resistance to drought, salinity, and disease. Recent interest in the use of annual species of Medicago as a cover and forage crop for use in sustainable agriculture systems has prompted both an evaluation of the diversity of germplasm that exists and the development of an annual Medicago spp. (medics) collection.
Medicago truncatula, M. polymorpha, and M. ciliaris are annual, self-fertilizing, and diploid and they grow naturally under different restrictive environmental conditions in the Mediterranean basin (Badri et al., 2016a, 2016b; Lesins and Lesins, 1979; Reid et al., 1989). M. truncatula has been chosen as a model for legumes because of its numerous advantages (Tang et al., 2014). It exhibits a high level of synteny with many cultivated legumes such as peas and perennial alfalfa (Young and Udvardi, 2009), thus allowing the transfer of knowledge on this model plant to legumes of economic interest (Brown et al., 2020).
These three Medicago species show different geographic distributions in Tunisia. M. truncatula is a ubiquitous species that grows in all bioclimatic niches (Arraouadi et al., 2009; Badri et al., 2007), while M. ciliaris ranges from humid to upper semiarid habitats (Badri et al., 2008) and M. polymorpha grows from humid to upper arid environments (Badri et al., 2016a; Haddoudi et al., 2021). They represent an important source of feed for livestock in Tunisia. However, these annual species are prone to fungal attacks that undoubtedly depreciate their nutritional quality. These pathogens include Phoma medicaginis and Cercospora sp. (Djébali, 2008).
Phoma is a highly polyphyletic genus with unclear species boundaries. It is a genus that has been described as the anamorphic phase of some species, of which the majority can be associated with the Didymellaceae ascomycetous family (Rai et al., 2014). Phoma is a pathogen for several types of legumes and causes black stem disease in Medicago species. The isolates of Phoma which cause disease in Medicago species belong to P. medicaginis and Ascochyta imperfecta (Ellwood et al., 2006). P. medicaginis is one of the most significant diseases on Medicago species in North America, South Africa, Europe, Australia and particularly Tunisia (Barbetti, 1983; Djébali, 2008; Graham et al., 1979; Lamprecht and Knox-Davies, 1984; Sampson and Western, 1941). Previous studies have shown variability in resistance to P. medicaginis in Medicago species (Barbetti et al., 2020; Djébali, 2013). This disease causes cankers on the stems and spots on the leaves. The attacked leaves turn yellow and eventually fall, causing complete defoliation of the plant during severe infection. The cankers on the stems grow and can reach the collar and the roots of the plant, causing their decline (Barbetti, 2007; Djébali, 2013). This disease spreads rapidly in cool and wet weather favorable for the development of conidial structures called pycnidia that grow on infected aerial and root parts (Djébali, 2013). The pycnidia can be preserved during winter on the dead plants and ensure contamination potential in spring.
Control of this disease can be achieved by using fungicides. Nevertheless, this control method is costly and has the drawback of development of resistant fungal strains. Extensive use of fungicides also poses risks to human health and the environment. Therefore, the development of alternative methods to control P. medicaginis, particularly by the selection of tolerant Medicago species, is a relevant way to control this disease in a crop rotation management strategy. Availability of disease-resistant accessions in the annual medic collection will enhance the utilization of these species in agro-ecosystems, and also may serve as a potential source of novel resistance genes amenable to incorporation into adapted genotypes. Sources of resistance in annual Medicago spp. to one of the most important indigenous foliar diseases of alfalfa, namely anthracnose, caused by Colletotrichum trifolii Bain & Essary, have been reported (Lamprecht, 1986; O’Neill and Bauchan, 2000).
The present work aims to analyze the responses to P. medicaginis Pm8 strain in a collection of M. truncatula, M. ciliaris and M. polymorpha inbred lines in order to use them as rotating crops to decrease disease pressure in the field and also to identify the source of resistance that can be used in future breeding programs.
Materials and Methods
Biological material and experimental conditions
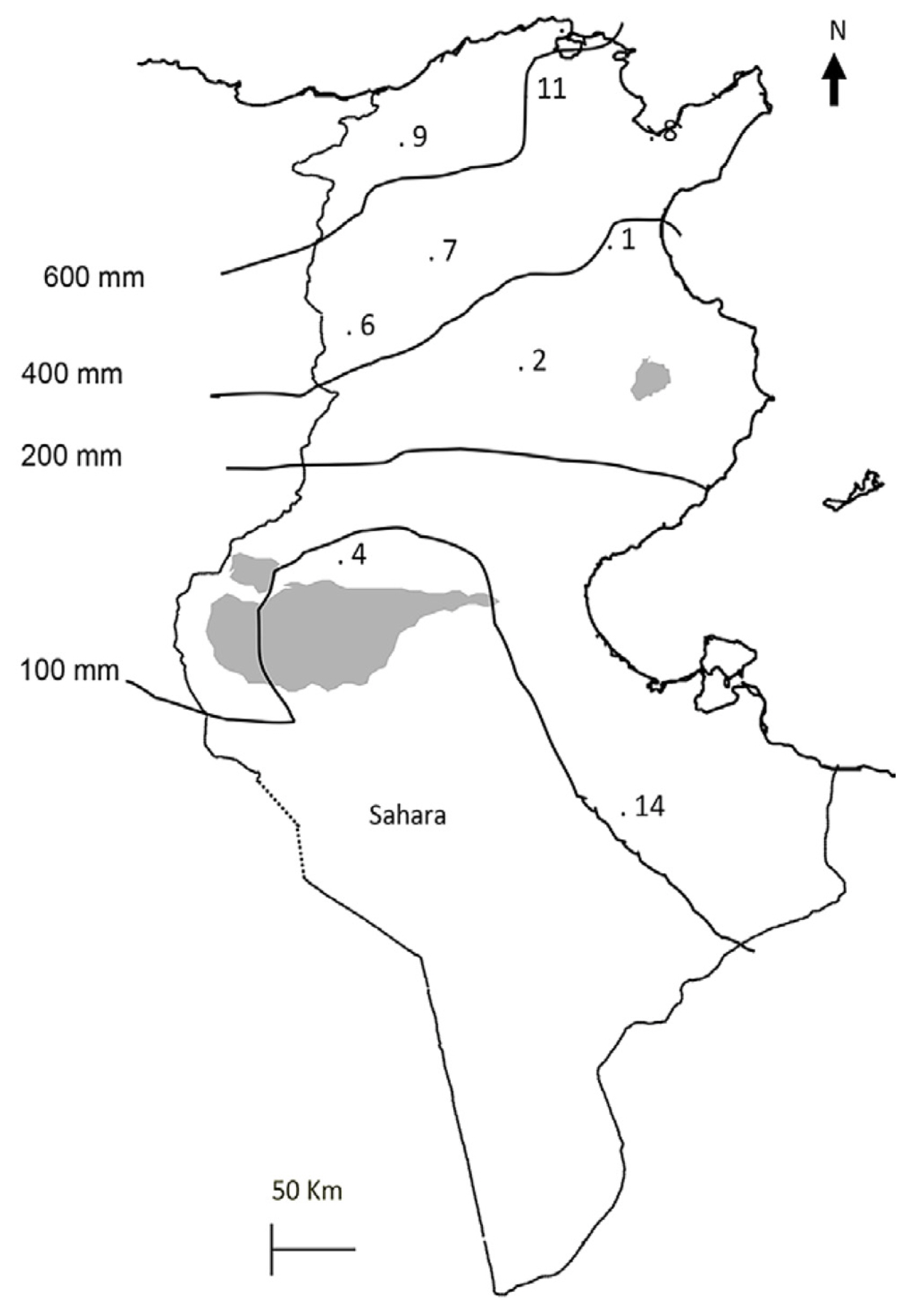
Forty-four Tunisian lines of M. truncatula, M. ciliaris, and M. polymorpha collected from different regions (Fig. 1, Supplementary Table 1) were used. In addition, the two reference lines Jemalong A17 and F83005.5 were also used in this study.
Seeds of lines were scarified mechanically using q600 glass paper. Then, the seeds were immersed in sodium hypochlorite 12% (NaOCl) for 3 min, then in 70% ethanol for 1 min. The seeds were then washed with sterile distilled water to remove the NaOCl and the ethanol. After imbibitions, the seeds were transferred onto agar medium (0.9% agar) in Petri dishes. The dishes were placed at 4°C overnight and then transferred to 25°C in the dark until the emergence of the radicle. Germinated seeds were transferred into square Petri dishes (12 cm × 12 cm) containing M medium (Bécard and Fortin, 1988). In order to avoid contact of the aerial part of the plants with the solid culture medium, the upper 1/3 of the medium was removed using a sterile scalpel. The Petri dishes were placed in a culture chamber at 25°C and a photoperiod of 16 h with a luminous intensity of 100 μE. The Petri dishes were placed vertically, with an inclination of 30° with the vertical plane to let the roots develop on the surface but not inside the medium (Supplementary Fig. 1).
The reference strain Pm8 of P. medicaginis (Djébali, 2013) was used for infection of Medicago lines. The fungal inoculum was prepared by scraping the surface of a three-week-old culture into sterile distilled water, which was subsequently adjusted to 106 conidia/ml. The Medicago seedlings were inoculated with 5 μl (106 conidia/ml) of the conidia suspension on the lower end of their main roots (3-5 mm of the root apex) 48 h after their transfer to M medium. The control plants were inoculated with 5 μl of sterile distilled water. Ten plants per line and treatment combination were used.
Trait measurement
Measurement of the parameters on the plants was carried out at harvest after 21 days post-inoculation. The measured parameters were the number of yellow cotyledons (YC), the number of green trifoliate leaves, yellow leaves, dead leaves (NDL), number of secondary roots (NSR), dead plants (NDP), root length (LR), and root browning length, and by measuring the fresh weight of leaves (FWL), roots (FWR), and fresh plant weight (PW). Determination of YC were performed visually. Estimation of the degree of infection of plants by P. medicaginis was made by calculating the percentage of browning on the roots (% Br), the number of pycnidia (fungal structures) on the roots (PycR) and on the leaves (PycL).
Data analyses
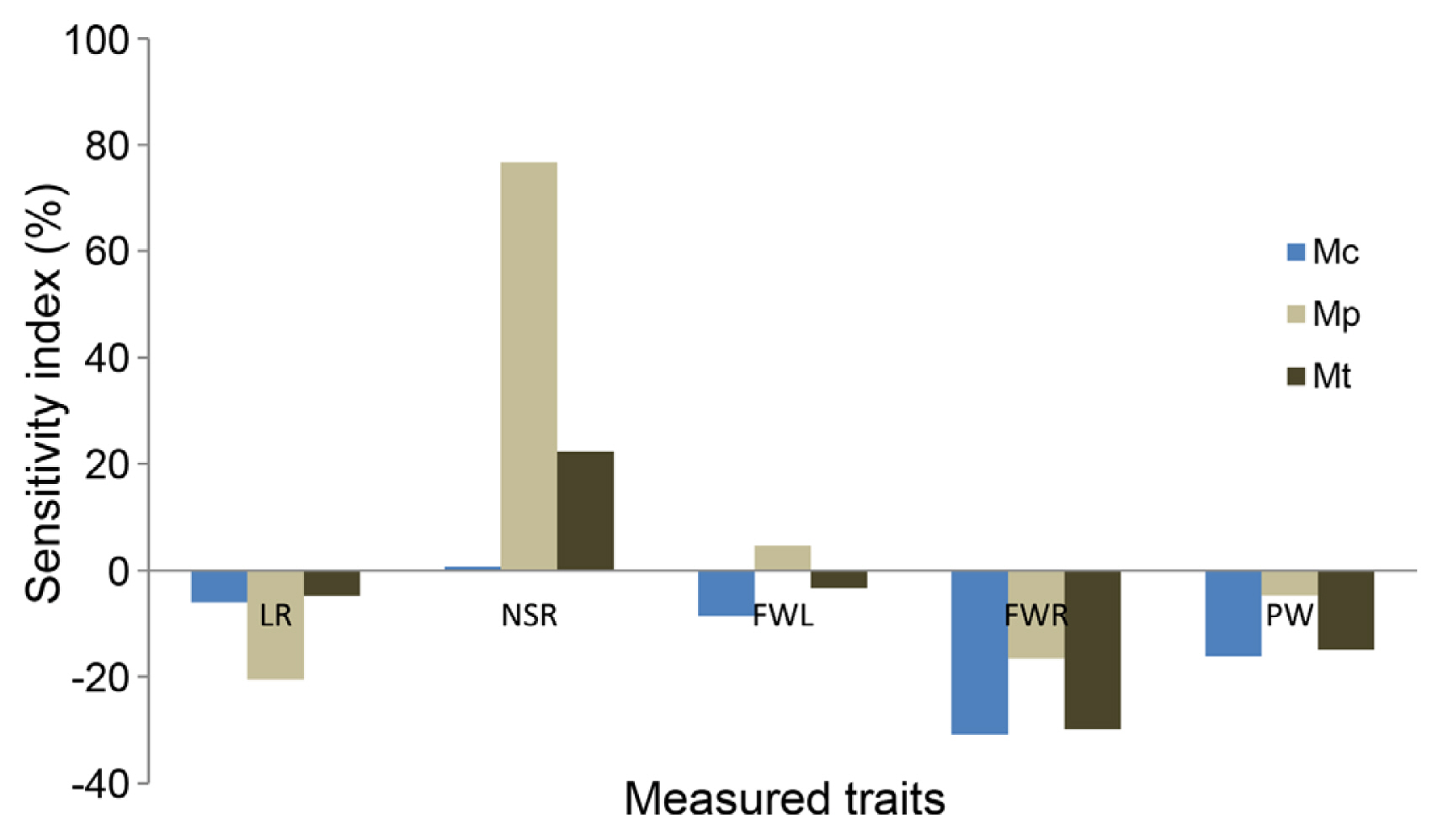
The results obtained were subjected to mono- and multivariate statistical analyzes. The analysis of the species effect, treatment effect, lines within the species interaction, species × treatment interaction, and line × treatment interaction for the measured traits was carried out using the generalized linear model (GLM) procedure contained in SPSS version software 20 (2014, IBM Corp., Armonk, NY, USA). Comparison of means of the measured parameters for the species and the lines was carried out using the Duncan’s multiple range test at P ≤ 5%. The sensitivity index (SI) in trait expression to Pm8 infection was estimated as follows (Arraouadi et al., 2011): SI = 100(S - C)/C, where S and C are the performances under Pm8 infection and control treatment, respectively.
Broad-sense heritability (H2) for each trait was estimated as H2 = Vg/(Vg + 1/k × Ve) (Badri et al., 2011), where Vg is the variance between lines, Ve is the environmental variance and k is the number of replicates per line.
The correlations between the measured parameters were estimated through calculation of the Pearson correlation coefficient (r). Principal component analysis (PCA) and hierarchical classification were performed based on the means of traits measured for studied lines under control treatment and fungal infection using XLSTAT software version 7.5 (Addinsoft, Long Island City, NY, USA). Discriminant analysis (DA) was performed using the means values for the lines groups.
Results
Morpho-physiological variation
The variation of measured parameters is justified based on the effects of species, lines nested within species, infection (treatment), species × treatment interaction, and lines nested within species × treatment interaction (Supplementary Table 2). The treatment factor exhibited the most important effect.
There is a variation of responses within and between the three species of Medicago to P. medicaginis Pm8 strain (Fig. 2).
In the control treatment, six (YC, LR, NSR, FWL, FWR, and PW) among the eight measured characters showed significant differences among the three studied species (Table 1). The highest values in aerial (FWL and PW) and root (LR and FWR) weights were registered for M. ciliaris followed by M. polymorpha and M. truncatula. Furthermore, ten among the 12 analyzed traits showed significant variation among the three analyzed species under Pm8 infection. Interestingly, the highest values of these traits were also noted for M. ciliaris followed by M. polymorpha and M. truncatula. Moreover, based on the SI, M. ciliaris was the least affected for the FWL (Fig. 3).
Most traits measured under both treatments showed that the highest levels of variation within species (CV) occurred for M. truncatula, followed by M. polymorpha and M. ciliaris (Table 1).
Broad-sense heritability (H2) of the measured traits ranged from 0.53 to 0.80 and from 0.20 to 0.55 under the control treatment and Pm8 infection, respectively. In the control treatment, high heritability (H2 > 0.40) was found for all measured traits. Furthermore, under Pm8 infection, high (H2 > 0.40) to moderate (0.2 ≤ H2 < 0.40) values of heritability were observed for analyzed traits.
In the control condition, among the 66 possible correlations between the measured parameters, 52 were significant and 26 correlations were positive (Supplementary Table 3). Negative correlations were recorded between the YC and NSR, FWL, and PW under both treatments. Among the 28 possible correlations for the traits measured in plants after infection with Pm8, 15 correlations were significant, of which 11 were positive. Comparison of the two correlation matrices showed specific correlations for the traits measured in plants after infection with Pm8.
PCA and cluster analysis
In the control treatment, the first two axes of the PCA explain 90.46% of the variability of the parameters measured for the three species of Medicago (Supplementary Table 4). Axis 1 is formed by NSR, FWL, FWR, and PW, while axis 2 is formed by the LR. Under Pm8 infection, the first two axes explain 62.93% of the variation of traits measured for the three studied species. The first axis is formed by NSR, FWL, FWR, and PW, while the second axis is formed by YC, PycL, NDL, and NDP.
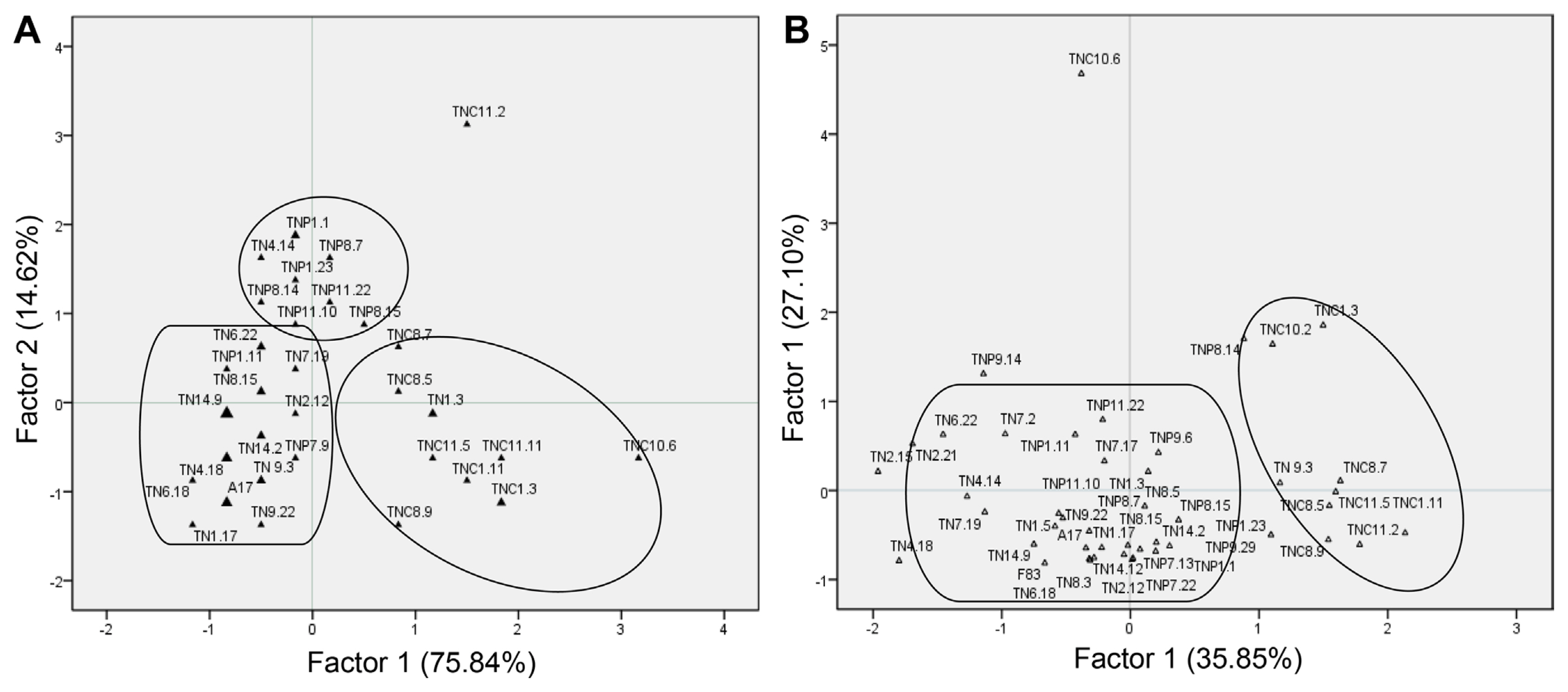
In the control treatment, the studied lines are structured into three groups according to their species of origin (Fig. 4). The first group contains the M. truncatula lines, the second group is formed by the M. polymorpha lines and a third group is constituted by the M. ciliaris lines. Under the Pm8 infection treatment, M. ciliaris lines formed a separate group distinct from M. truncatula and M. polymorpha lines which were grouped together.
Hierarchical classification (CHA) of the studied lines showed that they are classified into two groups under control treatment and Pm8 infection (Fig. 5). In the control treatment, the first group is formed by 32 lines of which 20 lines are from M. truncatula and 12 lines from M. polymorpha. The second group is composed of 14 lines of which 11 lines are from M. ciliaris, two lines from M. polymorpha and one line (TN1.3) from M. truncatula. Under Pm8 infection, the first group is constituted by 38 lines of which 22 lines are from M. truncatula, 17 lines from M. polymorpha, and one line (TNC10.6) from M. ciliaris. The second group is composed of eight lines from M. ciliaris.
DA showed that LR, NSR, FWL, FWR, and PW are the determinant characters in the classification of the lines in two groups under both treatments (Table 2). The lines from group 2, most of which belong to M. ciliaris, exhibited the highest values for these traits.
Discussion
Accessions and cultivars of both annual and perennial Medicago spp. exhibit a wide range of responses to disease, including P. medicaginis, Erysiphe pisi, and C. trifolii (Barbetti et al., 2020; Yaege and Stuteville, 2002). Spring black stem and leaf spot caused by P. medicaginis is a common and serious disease in Africa, America, and Europe. It infects both above and underground parts of the plants (Djébali, 2013). Plant productivity losses can be high while cultivars with acceptable levels of resistance are not available. Therefore, several researches are oriented towards the exploitation of the genetic diversity in annual Medicago species in order to understand the resistance mechanisms and to find new resistance genes against legume pathogens, including P. medicaginis (Djébali, 2008; Moussart et al., 2007; Pilet-Nayel et al., 2009).
The line × treatment interaction noted for the majority of the characters suggests the utility of this collection of lines for the identification and characterization of genetic determinants for resistance to P. medicaginis.
In this study, our results showed that most variation of measured parameters is explained by the infection treatment factor. Moreover, the significant line × treatment interaction argues in favor of the utility of this collection of lines for the characterization of the genetic basis of resistance to P. medicaginis. This suggests that the performance of lines has comparable trends for most of the measured traits. Similar trends were also obtained by Djébali (2013) in aggressiveness and host range of P. medicaginis isolated from Medicago species growing in Tunisia.
Tivoli et al. (2006) suggested that some degree of resistance to many diseases is already available in some annual Medicago spp. For example, variation in resistance to stem and leaf disease caused by P. medicaginis has been reported in annual Medicago spp. Andrew (1962) demonstrated that M. denticulata had much greater resistance to post-emergence by disease caused Pythium spp. than M. minima did. Therefore, the wide range of negative effects of P. medicaginis on the measured traits argues for the benefit of making an exhaustive selection for resistance to this disease in Medicago species.
Means comparison of analyzed characters showed that the highest values were observed for the lines of M. ciliaris under the control condition and under Pm8 infection. Most diversity of response to Pm8 within species was found for M. truncatula in comparison to M. polymorpha and M. ciliaris, suggesting some specific host-pathogen coevolution traits since Pm8 strain was originally isolated from M. truncatula. So, it is worthy to test this hypothesis by cross inoculating P. medicaginis strains isolated from these three Medicago species. Accordingly, Boerema et al. (1965) showed that black stem fungi isolated from alfalfa and red clover could attack both plant species, but the isolates were more pathogenic on the host from which they were isolated.
Most of the measured traits showed high to moderate heritability (H2) levels under both treatments suggesting that these characters could be used as good descriptors for the selection of the lines of annual Medicago species tolerant to P. medicaginis infection.
The application of correlation analyses between two or more characters is an important indication to understand how improvement in one character could cause simultaneous changes to other characters (Falconer and Mackay, 1996). Our results showed that correlations established between measured traits are affected by the infection treatment effect, where some correlations were noted when the plants are infected with the Pm8 strain. The three annual Medicago species distinctly differed for response to P. medicaginis. There were no genotypes with a highly effective resistance; so breeding for partial resistance to the Pm8 strain is likely to be achieved which provides a long-term resistance behavior for breeding varieties (Keller et al., 2000).
Results from PCA and cluster analysis of the studied Medicago species could be a useful guide for selection programs. The lines studied formed two different groups according to their resistance to P. medicaginis. Most lines showed a certain level of tolerance to the disease. Lines combining tolerance to this disease, identified in this study, and to drought stress identified in previous work (Badri et al., 2016b) will be selected and studied under these combined constraints simultaneously to select lines that cope with both stress since they coexist in the field conditions in Tunisia and several Mediterranean environments.
In conclusion, our results showed that most variation in the measured parameters is explained by the treatment factor. Moreover, the significant line × treatment interaction argues in favor of the utility of this collection of lines for the characterization of the genetic basis of Medicago resistance to P. medicaginis. Means comparison of traits showed that the highest values were found for the lines of M. ciliaris under both treatments. Correlations established between measured traits are affected by the treatment effect where some specific correlations are noted when the plants are infected with the Pm8 strain. Most diversity of resistance to Pm8 within species was found for M. truncatula. The strains studied formed two different groups according to their resistance to P. medicaginis. Most lines of M. ciliaris showed superior vigor in the presence of the disease. This species can be used as a good candidate in crop rotation strategy to decrease disease pressure in field and as a source of resistance genes.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement1
Supplement1 Print
Print







