Light and Electron Microscopy Studies Elucidating Mechanisms of Tomato Leaf Infection by Pseudocercospora fuligena
Article information
Abstract
The fungal pathogen Pseudocercospora fuligena, known to affect tomatoes in the tropics and subtropics, has been reported from temperate climates including the United States and Turkey in recent years. In this study, an isolate from fresh tomatoes and the disease it causes were characterized and infection mechanisms investigated. Macroscopically, both sides of tomato leaves show indistinct effuse patches but prolific production of fuliginous lesions is conspicuous on the abaxial side first but also on the adaxial side later on as infection progressed. Microscopically, fascicles of conidiophores (11–128 μm × 3.5–9 μm) arising from stromata and conidia with up to 12 septations were observed. Molecular characterization of the isolate revealed high homology (99.8%) to other P. fuligena isolated from tomatoes in Turkey. Out of the 10 media tested, P. fuligena grew significantly well and sporulated better on unsealed tomato oatmeal agar and carrot leaf decoction agar, both supplemented with CaCO3. Direct transfer of conidia from profusely sporulating lesions was the easiest and quickest method of isolation for in-vitro studies. Light and scanning electron microscopy on cleared and intact tomato leaves further confirmed stomatal penetration and egress as well as prevalence of primary and secondary infection hyphae. In situ, blocked stomatal aperture areas of 154, 401, and 2,043 μm2 were recorded at 7, 12, and 17 days after inoculation, respectively. With the recent expanded horizon of the pathosystem and its consequential impact, such studies will be useful for a proper diagnosis, identification and management of the disease on tomato worldwide.
Pseudocercospora fuligena (Roldan) Deighton, also synonymously called Cercospora fuligena (Roldan), causes black leaf mold (BLM) disease on tomato (Lycopersicon esulentum Mill.). The pathogen and the disease it causes were first described from the Philippines by Roldan (1938) and for long known as “Cercospora leaf mold” until it assumed the current taxonomic nomenclature after Deighton in 1976. P. fuligena is among the hyphomycetous fungi belonging to the group dothideales whose teleomorphs, if known, belong to Mycosphaerella within ascomycotina (Crous et al., 2004). However, several thousand anamorph species exist that lack known teleomorphs (Crous and Braun, 2003). An isolate used in this study, from a sample collected in Thailand, was grown in culture at the Institute of Plant Diseases and Protection in Germany, DNA extracted using internal transcribed spacer (ITS) primers and submitted to the GenBank. Separately, a diseased leaf sample was sent and stored in CBS culture #132017 and has been partially sequenced using 28S ribosomal RNA gene (GenBank accession no. JQ324953) and included in an extensive review of phylogenetic lineages in Pseudocercospora (Crous et al., 2013).
P. fuligena, which is known to be predominantly prevalent in tropical and subtropical countries (Halfeld-Vieira et al., 2006; Hartman and Wang, 1993; Wang et al., 1996) has recently been detected in Ohio (Subedi et al., 2015), North Carolina (Lookabaugh et al., 2018) after the last report from two counties in Florida (Blazquez and Alfieri, 1974). It is also of reported from Turkey in Europe (Gül et al., 2021). Such trends of detection in new geographic locations is alarming and calls for future studies investigating whether mixed infections exist or whether a shift in climatic conditions resulted in expanded horizon of the pathogen. Damage inflicted by the pathogen and the yield loss reported differs based on season and tomato varieties planted. Hartman et al. (1991) reported up to 32% of yield loss due to BLM on susceptible varieties without fungicide application in Taiwan. Mersha et al. (2014b) reported an average of 30.6% yield loss from two peak epidemic periods in Thailand.
The extent of yield loss across regions due to the expansion of P. fuligena in the United States, Europe, and potentially other places which otherwise were known to be affected by another leaf mold disease caused by Passalora fulva also synonymously known as Cladosporium fulvum remains to be studied. Despite a continued effort to increase productivity of tomato in open fields and protected culture in the United States and elsewhere, frequent detection of P. fuligena on tomatoes calls for more research and outreach endeavors on this pathosystem.
Mersha et al. (2014a) presented intensive investigation on monocyclic components such as incubation and latent periods of BLM based on wetness duration, temperature, and leaf age. In addition, Mersha and Hau (2009) published outcomes of comparative epidemics studies of BLM caused by P. fuligena and early blight caused by Alternaria solani in four different greenhouses. Furthermore, spatio-temporal dynamics of the disease under protected cultivation from two foci has been reported (Hau and Mersha, 2009).
The current study investigated the biology pathogenicity and infections mechanisms of P. fuligena in situ and in vitro as well as symptomatology of the BLM disease based on the isolate (GenBank accession no. JQ324953), the successive greenhouse studies under natural infection in Thailand and growth chamber studies in Germany. The study also includes recent reports of the pathosystem from the United States in North America and Turkey in Europe. The latter is implicated in concerns of growers, gardeners, educators, and Ag consultants as to whether there is a shift in the pathosystem complex of tomatoes in the United States or whether a mixed infection exists between the two pathogens mentioned above. This study will compliment recent effort in sequencing the genome of P. fuligena (Zaccaron and Stergiopoulos, 2020) and future research arising from the aforementioned concerns. Outcomes of the current study are timely and useful to contrast pathogenesis by the two pathogens and symptomatology among the two leaf mold diseases thereby highlight the implications of the appropriate preventive and curative strategies.
Materials and Methods
Macroscopic features, isolation, and culture morphology
Symptoms and signs of BLM disease were characterized based on digital pictures captured from the adaxial and abaxial leaf surfaces of the tomato cultivar FMMT260. Three methods of isolation, namely direct conidial transfer (DCT), conidial suspension transfer (CST), and lesion transfer from advancing margin (LT) were compared on tomato oatmeal agar (TOA) prepared according to Hartman et al. (1991) and amended with CaCO3. For DCT, 6-mm-sized lesion with profuse sporulation was aseptically slid at four lines running across the diameter of a 9-cm diameter Petri dish. For CST, a drop of a P. fuligena conidial suspension (2 × 104/ml) was dropped at four corners and spread to form four lines using sterile inoculation loop over the 9-cm diameter Petri dish; For LT, four profusely sporulating lesions were aseptically placed at four corners across the 9-cm diameter Petri dish. The experiment, with the three treatments (DCT, CST, and LT), was arranged in a completely randomized design with 10 replications repeated once. Growth of a colony and extent of contamination was recorded 72 h after incubation at 28°C under dark on the basis on a 0–3 scale. Contamination was visually scored in a scale of 0 (0%; clean, no contamination traced visually), 1 (1–20%; slightly contaminated), 2 (21–50%; intermediate contamination), or 3 (≥51%; extensively or completely contaminated); in vitro growth of P. fuligena was also visually rated on a scale of 0 (1–10%; poor growth), 1 (11–25%; low growth), 2 (26–60%; intermediate growth), or 3 (>60%; extensive high growth, visible).
Furthermore, morphology and growth of P. fuligena was studied in the following 10 growth media, with or without sealing; biomalt agar (BMA), 15 g biomalt and 10 g agar powder in 1 liter of distilled water; carrot leaf decoction agar (CLDA), 300 g finely ground carrot leaf, boiled in 500 ml distilled water, steamed for 1 h and strained through a double layer of cheese cloth, filtrate mixed with 500 ml distilled water in which 12 g agar has been dissolved; carrot leaf oatmeal agar (CLOA), 50 g oatmeal boiled in 500 ml for 20 min and filtered, 50 ml carrot leaf juice, 20 g agar mixed and distilled water added onto the filtrate to adjust the volume to 1 liter; malt extract agar (MEA), 50 g of dehydrated powder (NutriSelect, Merck KGaA, Darmstadt, Germany) mixed in 1 liter distilled water and autoclaved at 121ºC for 15 min; oatmeal agar (OMA), 72.5 g of dehydrated powder (NutriSelect, Merck KGaA) mixed in 1 liter distilled water and autoclaved at 121ºC for 15 min; potato carrot agar (PCA), 15 g potato, scrubbed and diced, 15 g carrot, peeled and diced, all boiled in 500 ml distilled water and sieved, 20 g agar added, volume adjusted to 1 liter by adding distilled water and mixture autoclaved at 121ºC for 20 min; potato dextrose agar (PDA), 39 g of PDA (Merck KGaA) mixed and in 1 liter of distilled water, autoclaved at 121ºC for 20 min; TOA, 50 g shredded tomato and 15 g oatmeal boiled separately each in 500 ml water, suspension sieved through two layers of cheese cloth and mixed, 15 g agar added, volume adjusted to 1 liter by distilled water, autoclaved 121ºC for 20 min. One g of CaCO3 was added to the mix just before autoclaving only for TOA amended medium for sporulation tests; vegetable agar (V8A), 200 ml V8 juice, 500 ml water, sieved and pH adjusted to 7–7.5 then 3 g CaCO3 and 10 g agar added and mixture volume adjusted to 1 liter by adding distilled water, autoclaved at 121ºC for 20 min; and water agar (WA), 10 g agar powder added to 1 liter distilled water, solution autoclaved at 121ºC for 20 min.
Profusely sporulating lesions (4-mm diameter) were cut aseptically and three pieces were gently pressed onto each media (point inoculation) at equally distant corner in a Petri dish. Each medium was replicated five times and incubated at 28°C in the dark. Colony growth (radial expansion in mm) was measured and extent of contamination was rated 7 days later using a 0–3 scale as shown in Table 1.
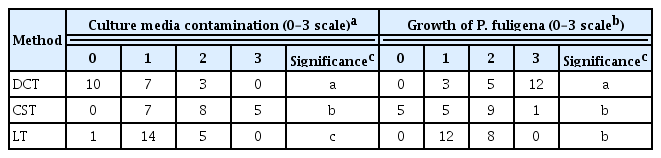
Contamination of culture media and in vitro growth of Pseudocercospora fuligena, rated on 0–3 scale and tallied for each of the three transfer methods, DCT, CST, or LT on a tomato oatmeal agar
Microscopic features: fungal structures
A profusely sporulating lesion was gently touched with a camelhair brush and the conidia, conidial fascicles, and conidiophores were dropped onto a drop of 0.5% acid-fuchsin on a clean glass slide for microscopic observations. Conidial shapes were determined, length and width measured and septations were counted using Olympus CX 40 light microscope (Tokyo, Japan) fitted with a camera and scaled using Motic Images Plus (Motic, Dongguan, China). In separate twice repeated experiments, conidia of P. fuligena that were brushed from infected leaves were dropped onto PDA, TOA or water and germination was recorded after 24 h incubation at room temperature in 12–12 dark to light durations. In another study, conidial germination was tested on detached leaves that were placed in small tightly closed plastic containers in dry, moist, and wet conditions in a greenhouse.
Microscopic features: mode of penetration, infection, and egress
Stomatal composition of abaxial or adaxial side of terminal leaflets sampled from the 6th internode of 2-week-old tomato plants was counted using a simple light microscopy. In another group of same aged plants, both sides of leaflets were marked, inoculated with a conidial suspension of 2 × 104/ml, covered with black plastic bags and kept in the greenhouse under high humidity for 16 h. Inoculated (and fully expanded) leaves were then directly stained with drops of 0.05% aqueous acid fuchsin solution 48 to 96 hours after inoculation (HAI). Stained leaflet section (6 mm) was randomly cut with a cork borer and infection mechanisms observed using stereomicroscope and later scanning electron microscopy (SEM) (Hitachi 3400N-SEM, Tokyo, Japan). For SEM, samples were prepared following the modified methods of Babu et al. (2002) and Blodgett and Swart (2002). Infection, sporulation, and progression of substomatal stroma were followed using techniques of two whole leaf clearing techniques by Bruzzese and Hasan (1983) and de Luna et al. (2002).
Molecular identification
Molecular characterization of an isolate was made from a diseased tomato leaf sampled from Thailand and cultured at the University of Hannover, Institute of Plant Diseases and Plant Protection in Germany. DNA was extracted from an axenic culture isolated from heavily BLM infected leaf using Chelex 100 method (Wichura, 2007). The complete ITS region was amplified using the universal ITS1 and ITS4 primers (White et al., 1990) and sequenced by MWG Biotech AG (Germany). DNA sequences were further aligned with the profile mode of Clustal X 1.81 (Thompson et al., 1997) and submitted to GenBank. A separate set of leaf sample was directly sent to the Centraalbureau voor Schimmelcultures (CBS) in The Netherlands (P. W. Crous, personal communication). For the latter, BLM symptomatic tomato leaf with profuse sporulation was collected, immersed in 70% ethanol for 15 s and then gently rinsed in two sets of distilled water, each lasting for a minute. Leaf sample was then blotted dry on sterile filter papers, pressed well and sent to CBS for molecular characterization.
Data collection and analyses
Quantitative data from the radial colony growth was analyzed using ANOVA (SAS version 9.4, SAS Institute Inc., Cary, NC, USA) and treatment means were separated using Tukey’s test at P < 0.05. The exact Wilcoxon test was used for the analyses of the frequency counts based on scales (Mehta and Patel, 2001). All regression analyses were made using Sigma Plot 12.0 (Systat Software Inc., CA, USA).
Results
Macroscopic and microscopic features. Initially, small, pale brown to yellow effuse patches were seen on both the upper and lower leaf surfaces of tomato leaves of the cultivar FMTT260 (Fig. 1). Within a day or two, depending on weather favorability, lesions on the lower leaf surface turned gray and then black (fuliginous) as P. fuligena, the causative agent of BLM sporulated (Fig. 1B and C). Sporulation and the fuliginous appearance were predominant but not exclusively hypophylous since the adaxial side also showed these signs when there is a prolific sporulation. Lesions coalesced as infection progressed leading to a complete drying of the leaf. However, leaves always remained hanging on the stem and hence no visible defoliation resulted by the disease unless mechanically disturbed by shaking.

Symptoms and signs of black leaf mold (Pseudocercospora fuligena) disease as viewed from the adaxial (A) and abaxial (B, C) leaf surfaces of tomato cultivar FMTT260.
Microscopic observations of conidia revealed a subhyaline to pale olivaceous, obclavate to cylindrical, attenuated tip, long obconic to long obconically truncated base, with slight constrictions at the septa, straight to mildly curved and unthickened scars (Fig. 2). A 0.5% aqueous fuchsin solution resulted in a better contrast of conidial structures within few minutes of staining (Fig. 2A). Germ tube that is exposed to 12 h of moisture (Fig. 2B) and fasciculate conidiophores (Fig. 2C) too were vividly stained using this solution. Average conidial dimensions recorded in this study were 11 to 128 μm in length and 3.5 to 9.0 μm width. More than 84.4% of conidia measured were 21 to 80 μm in length with an average length of 45.9 μm (n = 192). Septation per conidium ranged from 2 up to 12 and showed a high linear correlation (r = 0.9, P = 0.002) with conidial length. Conidiophores were typically loosely fasciculate, pale olivacious to pale brown in color, straight to sinuous, tip rounded or truncated, sometimes once geniculate and not branched.

Microscopic features of Pseudocercospora fuligena in fuchsin stained mount: (A) conidia, (B) germinating conidia, and (C) fasciculate conidiophores.
Isolation and morphology on culture medium
DCT proved to be the most efficient and expedient method of isolating P. fuligena, with a significantly lower rate of contamination but fast growth compared to the other two methods (Table 1). In addition, inoculum was widely spread on the growth media using zigzag or cross streaking thus found to be by far the most practical and efficient in more prolific growth of the fungus within a short time compared to point inoculations. Consequently, DCT was used in many experiments of this study including in vitro isolation and inoculations of P. fuligena.
P. fuligena grew significantly better on TOA and CLDA supplemented with calcium carbonate and when incubated in non-sealed Petri dishes (Table 2). Respective radial growth (mm) in a non-sealed Petri dish was 13.6 in TOA and 13.1 in CLDA 7 days after inoculation (DAI) which was significantly higher compared to others (Table 2). The least growth was recorded on OMA, PCA, V8A, and WA (Table 2). Better growth was also seen in non-sealed Petri dishes of TOA, CLDA, and MEA when contrasted to their counterparts which were wrapped and sealed (S) with parafilm.
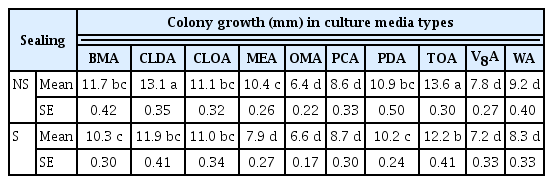
Colony growth of Pseudocercospora fuligena in terms of diameter expansion in millimeter following point inoculation of profusely sporulating lesions in 10 different culture media with (S) and without (NS) sealing Petri dishes
Molecular characterization
Polymerase chain reaction product sequenced from the DNA extraction of the fungal pathogen cultured on TOA revealed a 470 bp which later was submitted to GenBank (accession no. OP984819) showed >99.5% homology to P. fuligena isolates from tomatoes in Turkey (GenBank accession no. MT828360), in Korea (GenBank accession no. JX290079) and two submissions from the same study area in Thailand (GenBank accession nos. GU214675 and GU060636). Pathogenicity was confirmed twice on three varieties (King Kong 2, Lizzy, and FMTT260) in a growth chamber study and the same pathogen was reisolated from inoculated plants. Non-inoculated plants were sprayed with distilled water and none of them showed BLM symptom. Outcome of the molecular characterization of the sample that was sent to the fungal biodiversity center in The Netherlands was also published as part of an extensive phylogenetic lineages study in Pseudocercospora (Crous et al. 2013).
Penetration and egress of P. fuligena
Stomata of tomato leaves including the subsidiary and guard cells were easily visible on both leaf surfaces of the intact (Fig. 3A) and cleared (Fig. 3B) leaves of FMTT260 viewed through light microscopy and intact leaves viewed at 500× magnifications using SEM (Fig. 3C). Though tomato plants under natural light situation are known to be amphistomatous, the abaxial surface was found to be more stomatophorous, with 148.4 ± 8.3 stomata/mm2 leaf surface, and hairy than the adaxial surface which has only 80.2 ± 7.6 stomata/mm2 leaf surface and often with a thicker cuticle. Microscopic observation of direct stomatal penetration of P. fuligena was possible through simple staining by 0.05% aqueous acid fuchsin solution. On intact inoculated leaves, primary infection hyphae were clearly discernible 48 HAI (Fig. 3D). Further spread of the pathogen by forming secondary infection hyphae was seen on detached leaves after leaf clearing (Fig. 3E).

Subsidiary cells, guard cells and stomata of a tomato leaf as seen through light microscopy of intact (A) and cleared (B) leaves and after scanning electron microscopy (C) and infection mechanisms of Pseudocercospora fuligena in terms of penetration through stomata of intact leaf (D) and dispersal through secondary hyphae on cleared leaf (E).
Leaf clearing and dyeing methods used in this study were useful in elucidating the mode of egress and progression of infection in situ by P. fuligena. Observation from light and electron microscopy confirmed stomatal mode of egress (Fig. 4). The earliest symptomatic appearances from artificial inoculations started about 6–8 DAI under optimal conditions in the greenhouse. The progressive incremental of stomatal blockage was clearly discernible from observations made at 7, 12, and 17 DAI on cleared but fuchsin stained leaves as seen through a light and electron microscopy (Fig. 4). The area (μm2), diameter (μm), and perimeter (μm) of these localized infection areas were significantly different among the three-time intervals and the progress of in situ infection during the first 17 days fitted best to two parametric exponential growth function with R2 > 0.81 (data not presented).

Pseudocercospora fuligena infection and fascicle formation through stomata as shown through contrasting of cleared leaves using light microscopy (A–C) and scanning electron microscopy (D–F) at 7 (A, D), 12 (B, E), and 17 (C, F) days after inoculation.
Microscopic observation of inoculated leaves 21 DAI showed the extent of enlargement of each fasciculate conidioma and also the conidia which were released to serve the dispersing the pathogen. Diameter of the area covered with conidiophores was 98 μm to 263 μm as it was observed from the light microscopy. The conidiogenesis study revealed the formation of conidioma and egress as observed through the SEM too (Fig. 4). The emergence of early conidiogenous cells through the stoma guard cells (Fig. 4A and D), the formation of the fasciculate conidiophores (Fig. 4B and E) and later sporulation (Fig. 4C and F) were evident.
Discussion
Macroscopic observations of BLM as well as the characteristic features of P. fuligena (Roldan) Deighton, from natural and artificial inoculations in this study, corroborated with earlier published studies that contrasted it with tomato leaf mold caused by Passalora fulva also synonymously known as Cladosporium fulvum Cooke (Syn. Fulvia fulva (Cooke)). Effuse patches of the lesion and amphigenous fruiting of the P. fuligena isolate from this study, was also noted by Deighton (1976) as one of the features distinguishing BLM from leaf mold caused by Cladosporium fulvum. Secondly, symptoms of the tomato leaf mold caused by Passalora fulva appears as olive-green to grayish purple in color but BLM caused by P. fuligena is initially white and turns gray to black (fuliginous) as infection progresses (Hartman et al., 1991; Sherf and MacNab, 1986). Nonetheless, initial lesion appearances of BLM caused by P. fuligena and leaf mold caused by Passalora fulva are still very similar in that leaf lesions of both turn pale green to yellow on the upper leaf surface and both fungi sporulate profusely on the lower leaf surfaces (Sherf and MacNab, 1986). In case of BLM caused by P. fuligena, lesions coalesced and leaves rolled upward and died prematurely at times of heavy infection with many leaves remain hanging on the plant with a soot-covered appearance (Wang et al., 1996). Our study also corroborated this whereas Hartman et al. (1991) reported premature defoliation due to BLM.
In situ microscopic observations of infection progression from cleared leaves at 7, 12 and 17 DAI in this research is also supported by similar studies in the past. For instance, typical symptoms of BLM did not appear until 10–14 DAI in field trials (Hartman and Wang, 1992) but lesions were visible 6 DAI and matured conidiophores and conidia 12 DAI in controlled conditions when incubated at 28°C (Wang et al., 1996). This reiterates the critical role of early preventive strategies to avoid pathogen infection or a diligent and timely intervention of curative options before the pathogen advances to completely block the photosynthetic apparatus of the tomato plant.
Microscopically, however, distinct morphology of conidia clearly distinguishes these two leaf molds as also stated by Hartman et al. (1991) and Wang et al. (1996). Passalora fulva is known to produce asexual conidia on clustered or single, tall dark, upright, variously branched conidiophores (Joosten and de Wit, 1999; Thomma et al., 2005; Ulloa and Hanlin, 2000) near the apex unlike the fasiculate types of P. fuligena. Fungal structures of P. fuligena from this research corroborated with published morphological variations. For instance, dimensions of conidia (length × width) ranged between 25–70 μm × 3.6–5 μm (Blazquez and Alfieri, 1974), 20–90 × 2.5–4 μm with 2–9 septations (Hsieh and Goh, 1990), 9–137 μm × 3.5–6.1 μm with 2–27 septations (Hartman et al., 1991), and 29–110 μm × 2.5–5.0 μm (Halfeld-Vieira et al., 2006). Our observations (11–128 μm × 3.5–9 μm with up to 12 septations) were close to all, particularly to the report from Taiwan (Hsieh and Goh, 1990).
Considering the slow growth of P. fuligena DCT from a profusely sporulating lesion was found to be efficient and less prone to contamination than the other two methods tested in this research, i.e., CST and point inoculation using LT. This information will be useful for future studies since establishment of the disease by artificial inoculation is essential component of many epidemiological studies and in turn for developing sound integrated disease management practices. The point inoculations from 6-mm diameter pure culture pieces in this study doubled in growth after 7 days of incubation at 28°C on TOA in the dark. But enhanced growth of P. fuligena was achieved through zigzag and radial spreading of point inoculations resulting in 104 to 105 propagules per Petri dish per 100 ml deionized water 14 days after incubation.
CLDA, a medium recommended for sporulation of Cercospora species (Kilpatrick and Johnson, 1956), and TOA, a medium that is widely used in World Vegetable Center, both supplemented with CaCO3, were the two culture media that consistently resulted in a significantly better growth of P. fuligena in this study followed by BMA, CLOA, PDA and malt extract agar. Earlier works of Hartman et al. (1991) have also reported comparable results and recommended the use of TOA. Interestingly, P. fuligena grew better in all non-sealed than sealed Petri dishes. Accordingly, colony grew significantly higher in non-sealed CLDA, TOA, and MEA with respective increased growth of 9.2%, 10.3%, and 24.0% in non-sealed compared to sealed Petri dishes.
For light microscopy, the whole leaf clearing method following de Luna et al. (2002) was preferred for its simplicity and omission of intermediary transfers saving the time to complete the procedure. In this method, cut pieces were immersed in vials containing 70 parts ethanol and 30 parts glacial acetic acid. It only took 30 min to stain the fungal structures in 0.05% acid fuchsin solution. After dyeing, fasciculate conidiophores were clearly visible with light microscopy. Though primary and secondary infection hyphae were observed, no cross-sectional study undertaken in this study to detect the internal hyphae, which according to Babu et al. (2002) develops inside leaf tissue and produces stromata in the substomatal chambers. Though stage of growth, canopy strata and light saturation determines stomatal composition, nearly a double stomatal density was recorded on the lower leaf surface compared to the upper.
Passalora fulva is predominantly prevalent in temperate climates including the United States as also confirmed by a recent study that presented a study of 50 isolates from high tunnels in the northeast and Minnesota (Sudermann et al., 2022). Nevertheless, P. fuligena, which is known to be predominantly prevalent in tropical and subtropical climates has recently been detected from Ohio (Subedi et al., 2015) and North Carolina (Lookabaugh et al., 2018) in the United States and from Muğla district in Turkey (Gül et al., 2021). Such a trend is alarming and calls for more studies and investigations to determine whether mixed infections exist in the United States and other temperate regions and/or whether global trends such as climate change are playing part in this shift. A potential future spread of the disease across regions could also be curbed with restricted movement of tomatoes and alternate hosts (Wang et al., 1995) from areas where the disease is endemic to new geographic locations. Hartman et al. (1991) also stated the need for additional research on geographical distribution and importance of these two diseases in areas of tropical and subtropical countries because of similarity of the field symptoms.
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.