



Citrus is one of the most important fruit crops worldwide, including in the Jeju province of South Korea, where citrus has been cultivated for over 100 years. Citrus production in South Korea was 30,799 ha of farm households in 2021, with a total income of 1,027,131 million won (Foreign Agricultural Service, 2020). Depending on many factors such as climate change, control management, etc., citrus melanose and gummosis caused by Diaporthe citri have been one of the serious diseases in many citrus orchards on Jeju Island. The disease is one of the factors responsible for not only decreasing citrus production but also reducing income. Melanose caused a decrease in income of 8.5% due to the reduced quality of fruits in the commercial market, which was much higher than the damage caused by other diseases or pests in Korea (Hyun et al., 2013). Therefore, the management of citrus cultivation to maintain citrus productivity and profitability is very important to the economics of Jeju Island.
D. citri is a fungus belonging to Ascomyceae (asexual stage: Phomopsis citri) and it produces the small black lesions raised pustules on the leaves, twigs, and fruits (Timmer et al., 1988) and produces inoculum in dead twigs as a saprophyte (Mondal et al., 2007). They have three types of conidia, alpha (α), beta (β), and gamma (γ). The α-conidia are hyaline, unicellular, fusiform to ellipsoidal and sized av. 6.7 × 2.5 μm. The β-conidia are hyaline, unicellular, filiform, curved, often strongly hooked and sized av. 22.1 × 1.0 μm (Kwon et al., 2003). The last γ-conidia are colorless, a multi-guttulata, fusiform to subcylindrical and sized av. 20.0 × 2.5 μm which are an intermediate between α- and β-conidial types (Chaisiri et al., 2022).
Melanose has been first identified in Jeju Island in 1971 (Baek et al., 1974) and the importance of control was not great until the late 1970s, but since the 1980s it has emerged as the most important disease due to the increase in cultivation areas, growing trees, and dense planting (Kwon et al., 2003). They also reported that increasing the risk of melanose epidemic was caused by increasing in citrus cultivation resulting in abundant trees on a farm. In Jeju Island, causing of melanose epidemic might be not only increased cultivation but also the spraying of chemical fungicides. Disease control of citrus trees is generally possible with chemical spraying 4 or 5 times a year (National Crop Pest Management System, 2020). In Korea, 95 items are used to prevent citrus melanose including the fungicides of benomyl, fluazinam, kresoxim-methyl, and prochloraz manganese (Rural Development Administration, System of Fungicide Security, 2020). However, controlling the disease with increased chemical pesticides also caused several negative effects such as the development of fungicide resistance of pathogen, deterioration of human health, and environmental risk (Gerhardson, 2002).
To establish a disinfectant resistance management system and reduce pesticide usage for the safety of agricultural production and income, monitoring of chemical resistance isolates of D. citri and screening of fungicides in the citrus production area of Jeju Island would be important issues. In our previous study, 40 D. citri isolates were identified using morphological and molecular biological methods on Jeju Island (Soe et al., 2023). This study aimed to monitor whether there is an outbreak of resistant isolate among these 40 D. citri against the commercial fungicides for the Jeju citrus production area by an in vitro test. Furthermore, the citrus leaves were bio-tested by inoculating them with resistant and susceptible isolates.
Materials and Methods
Fungal isolates
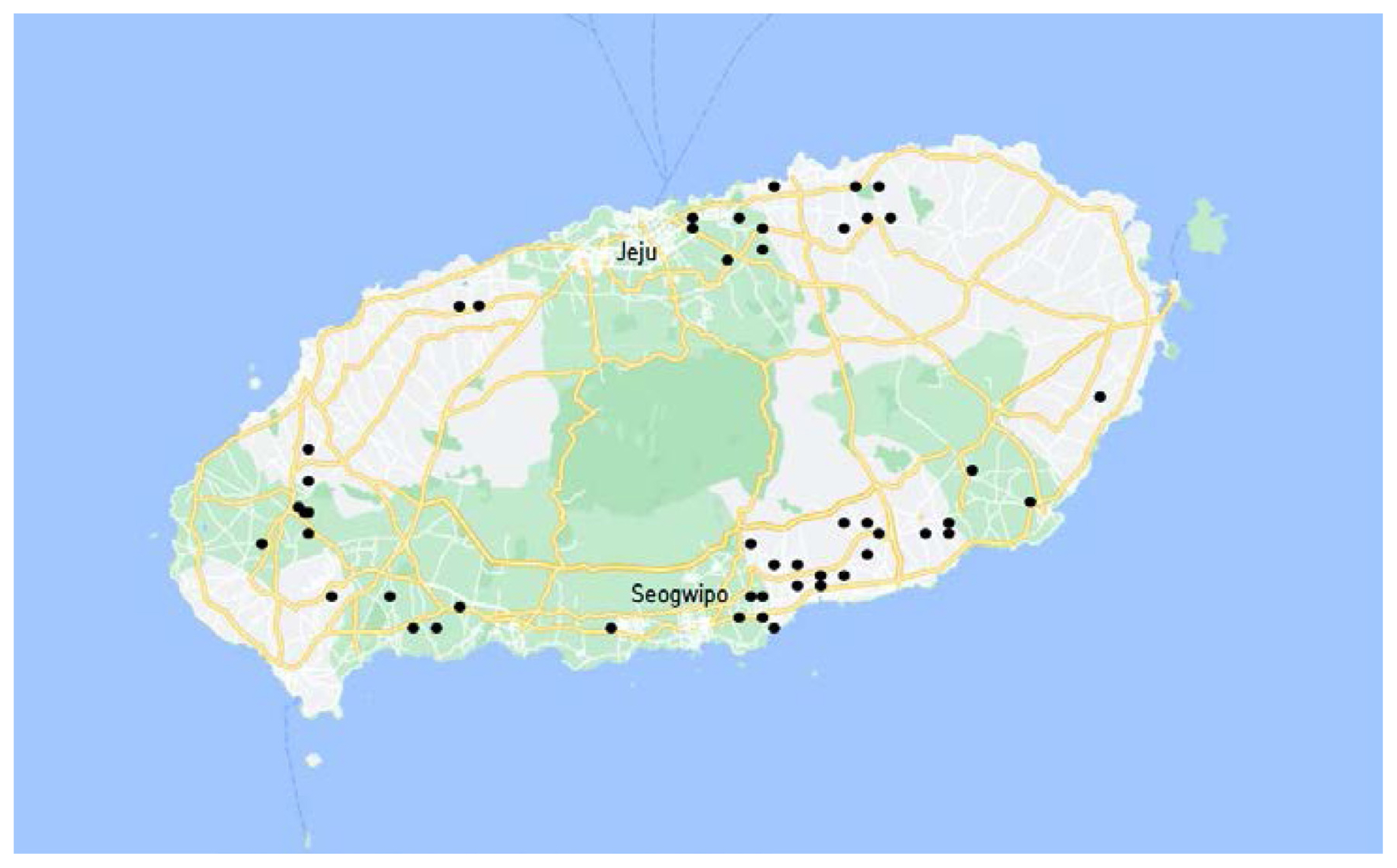
Forty D. citri isolates, which were collected from 49 citrus orchards from northeastern, southeastern, northwestern and southwestern regions of Jeju Island from March to April 2022 (Fig. 1), were used for chemical resistance experiment.
Evaluation of fungicide resistance
Four fungicides were used to evaluate chemical resistance in the isolates of citrus melanose (Table 1). To determine an effective dose for 50% inhibition (ED50) of the agrochemicals, firstly, D. citri KJ-14, which is a strain isolated in Jeju Island, was incubated on potato dextrose agar containing various concentrations of each chemical. The concentration inhibiting hyphal growth of the pre-tested isolate to 50% compared with that of on potato dextrose agar (PDA) without agrochemical was determined as a ‘criteria concentration’ for each agrochemical to test the ED50 of forty isolates. To determine for the first time whether the tested isolate is resistant or susceptible, it needed certain concentration of the chemicals, which was referred to as ‘criteria concentration’. Two lower (5 times and 25 times) and three higher concentrations (5 times, 25 times, and 125 times) than the criteria concentration of each agrochemical were used for amending the PDA media plates to test the chemical resistance of the fungal isolates. The fungal isolate inoculated on the PDA without agrochemical was used as a control. A total of 40 isolates of citrus melanose were inoculated on the PDA containing the four agrochemicals at seven concentrations with three replications, respectively.
The inoculated plates were incubated at 28°C under dark conditions until three weeks to get the fungal hyphae fully grown on control plates. To calculate the inhibition rate (%) of mycelium growth, the diameter of the isolated colony on PDA which was treated with fungicide and without fungicide was measured with a ruler. The mycelium growth inhibition percentage is calculated according to the following formula:
I (%), inhibition percentage; Dc, average diameter of the control colonies; Dt, average diameter of the treated colonies.
Determination of ED50
To represent the activity/dose relationship between agrochemical and fungal growth, a linear dosage-response line for the most effective concentration was drawn by plotting the average value of inhibition rate of susceptible fungal isolates and the following equation was used.
y, inhibition percentage; x, effective concentrations; a, a constant; b, the slope of the line
Bioassay evaluation
Young leaves of citrus, which were not hardened, were inoculated with representative chemical-resistant and susceptible D. citri isolates. The citrus leaves were soaked in 1% sodium hypochlorite solution (NaClO) for 30 s, followed by 70% ethanol for 30 s, then rinsed three times in sterile distilled water for 30 s. These leaves were laid in a Petri dish (ø = 90 mm) with sterilized filter paper. The end of the petiole was wrapped with cotton wool soaked with sterilized water. These citrus leaves were pre-sprayed with kresoxim-methyl at 0.5 ml/L or benomyl at 0.65 g/l for 3 h of which chemical resistance were found by some isolates of D. citri. As a negative control unsprayed leaves were used. Inoculum suspension adjusted to 1 × 106 conidia/ml of D. citri. After drying leaves for 3 h, the inoculum added with 0.01% Tween 20 (BIOSESANG Co., Ltd., Yongin, Korea) was dropped onto the left side of leaves by resistant isolates and the right side of leaves by susceptible isolates with 3 points every 20 μl each followed by incubation in an incubator (DA MIL-2500, Dong-A, Siheung, Korea) at 25°C until clear symptoms. Diameters of infected sites showing symptoms of melanose were measured with a ruler. Experiments were done with three replications in one leave and every treatment contained six leaves.
Statistical analysis
The data gathered from the chemical resistance experiment and inoculation experiment was analyzed using Statistical Tool for Agricultural Research (STAR) version 2.0.1 (IRRI-STAR, 2013). Treatment means were compared by using Tukey’s honestly significant difference test and least significant difference test, respectively.
Results
Criteria concentration
The criteria concentrations were obtained for benomyl as 0.25 μg/ml, fluazinam as 0.061 μg/ml, prochloraz manganese as 1.45 μg/ml, and kresoxim-methyl as 0.0013 μl/ml by comparing the diameter of the mycelium grown on a pre-tested isolate between the PDA without chemical and the PDA with chemical.
Evaluation of fungicide resistance
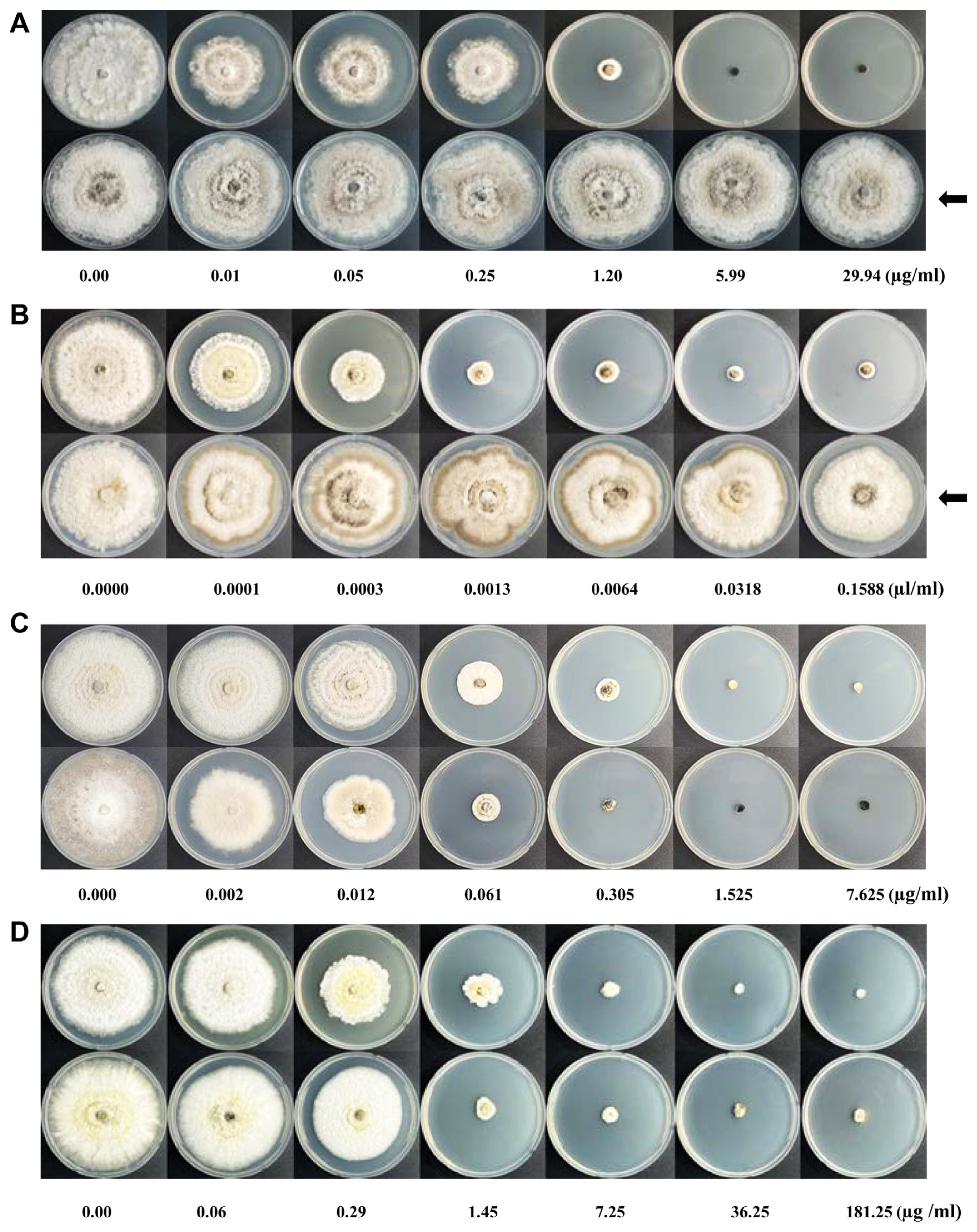
With the escalating concentrations of the benomyl, the mycelial growth diameter of the representative fungal isolates, except NEL21-2, was significantly different (P < 0.001) (Supplementary Table 1). On the higher fungicide concentrations, benomyl inhibited strongly the hyphal growth in which no fungal growth was observed in the most fungal isolates except NEL21-2 (Figs. 2A and 3A, Supplementary Table 1). To calculate the ED50, the mycelium diameters in the concentrations of 0.01 to 1.2 μg/ml were selected but with neglecting the resistant isolate NEL21-2 (Fig. 3B). Remarkably, with benomyl fungicide, only one isolate NEL21-2 was identified as a resistant isolate against benomyl of which hyphae grew well even on the PDA medium containing the highest concentration of benomyl (Figs. 2A and 3A). Also, the mycelial growth of NEL21-2 was not significantly different (P ≥ 0.05) within the tested concentrations of fungicide. Among the susceptible fungal isolates, NEL 6-1 seemed to be more susceptible to the lower concentration of benomyl than the rest fungal isolates (Figs. 2A and 3A).
The mycelial growth diameter of the representative fungal isolates with fluazinam and prochloraz manganese was also found as highly significantly different (P < 0.001) with the escalating concentrations of both fungicides (Supplementary Tables 2 and 3). Fluazinam (Figs. 2C and 3C) and prochloraz manganese (Figs. 2D and 3E) also caused the hyphal growth inhibition and no growth or the slowest growth on the higher concentrations. The diameter of mycelial growth of 0.002 to 0.305 μg/ml of fluazinam and 0.06 to 7.25 μg/ml of prochloraz manganese were assessed for the calculation of ED50, respectively (Fig. 3D and F). However, no resistant fungal isolates were found among them against both chemicals (Figs. 2C and D, 3C and E).
Unlikely, the fungal growth on the medium containing kresoxim-methyl was different from those on the medium with other three fungicides. All fungal isolates could grow at the highest concentration of kresoxim-methyl (Figs. 2B and 3G, Supplementary Table 4) whereas most fungal isolates didn’t grow at that concentration of the three chemicals (Fig. 2A, C and D). Among the 40 fungal isolates, 12 fungal isolates were selected as resistant isolates against kresoxim-methy which could not inhibit the fungal growth to more than 50% (Fig. 3G), regarding as resistant isolates in this study. The mycelial growth diameter in the concentrations of 0.0001 to 0.0064 μl/ml of kresoxim-methy was selected to calculate the ED50 by neglecting the resistant isolates (Fig. 2H). Among the resistant fungal isolates, SEL 19-1 was the most resistant (Figs. 2B and 3G) and SEB 3-1 and SEB 3-2 seemed to be more susceptible to kresoxim-methyl (Figs. 2B and 3G).
Determination of ED50
The data relating to the inhibition rate (%) of susceptible fungal isolates (Supplementary Tables 5 and 6) depending on the four concentrations by excluding the highest two concentrations of each agrochemical were subjected to linear regression analysis, in which r2 value was calculated as 0.99 for benomyl, 0.91 for fluazinam, 0.81 for prochloraz manganese and 0.71 for kresoxim-methyl, respectively. ED50 value for each fungicide was calculated using the regression equation as 0.66 μg/ml for benomyl, 0.18 μg/ml for fluazinam, 2.95 μg/ml for prochloraz manganese and 0.0045 μl/ml for kresoxim-methyl, respectively, against the susceptible D. citri (Fig. 3B, D, F and H).
Bioassay evaluation
As a couple of sets of resistant and susceptible isolates, NEL7-2 (resistant)/NEB7-3(susceptible) for kresoxim-methyl, and NEL21-2 (resistant)/NEL6-1(susceptible) for benomyl were used for bioassay evaluation. The results of the resistance in vitro tests of NEL7-2 to kresoxim-methyl and NEL21-2 to benomyl were coincide with the results of the inoculation tests in vivo. On the chemically unsprayed leaves, apparent diseased lesions appeared after inoculation with all isolates of D. citri used in the in vivo test (Fig. 4A and C). However, a couple of sets of comparisons of resistant and susceptible isolates to kresoxim-methyl showed that resistant D. citri isolate NEL7-2 had higher disease severity compared to susceptible isolate NEB7-3 (Fig. 4B). Furthermore, the average diameter of diseased lesions in resistant isolate NEL7-2 was significantly larger than that in susceptible isolate NEB7-3 (Fig. 4E). Moreover, the results of disease severity (Fig. 4D) and the average diameter of diseased lesions (Fig. 4F) with benomyl pointed out the same trends with kresoxim-methyl.
Discussion
In this study, four chemicals were used to screen the chemical resistance of isolated D. citri in order to establish a disinfectant resistance management system and reduce pesticide usage for the safety of agricultural production in Jeju Island. The results of Liu et al. (2023) showed that both quinone outside inhibitor fungicides kresoxim-methyl and trifloxystrobin inhibited conidial germination of D. citri up to 100% at 0.1 μg/ml, and kresoxim-methyl significantly inhibited the infection process compared to the control treatment in vivo test of detached fruit with 0.1 g/l. But our findings had 28 sensitive isolates to kresoxim-methyl and the average ED50 for the susceptible was 0.0045 μl/ml. And 12 isolates against kresoxim-methyl which could not inhibit the fungal growth to more than 50% at the highest concentration of 0.1588 μl/ml (Fig. 3G). The results of resistance in vitro tests have coincided with those of some fungus inoculation tests in vivo at 0.5 ml/l (Fig. 4B and E). There was a report in which strobilurins fungicides were not as effective in controlling melanose (Zitko and Timmer, 1998).
The sensitivity and resistance of D. citri to kresoxim-methyl may be in line with some of the following findings. Like all strobilurins, kresoxim-methyl takes action as a specific inhibitor of respiration by binding to the center Qp of cytochrome b (Anke, 1995; Baldwin et al.,1996; Becker et al., 1981; Clough et al., 1995; Mizutani et al., 1995; Sauter et al., 1995; Von Jagow and Link, 1986). The site-specific mode of action of strobilurin fungicides raises the possibility of resistance development (Köller, 1991). In addition to target site mutations, strobilurin sensitivities can also be reduced by inducing alternative respiration in response to the action of respiration inhibitors such as strobilurins (Day et al., 1995). This alternative pathway, which is active in the presence of alternative oxidase (Li et al., 1996), has been identified as the cause of fungal mycelia’s low strobilurin sensitivities (Hayashi et al., 1996; Mizutani et al., 1995, 1996; Olaya et al., 1998; Shirane et al., 1995). This mechanism of mutational strobilurin resistance or natural tolerance has not been reported for any phytopathogenic fungus. For Venturia inaequalis, the amino acid sequence determined for the cytochrome b target site indicated full sensitivity to strobilurins (Zheng and Köller, 1997).
Benomyl also showed resistance as one isolate NEL21-2 whose mycelia growth inhibition was 0.98% at the highest concentration (29.94 μg/ml) and also was not effective at all tested concentrations of fungicide (Fig. 3A). Results from in vivo testing of resistant NEL21-2 and susceptible NEL6-1 at 0.65 g/l (Fig. 4D and F) were in line with results from in vitro tests (Figs. 2A and 3A). But most of the isolates except NEL21-2 were found as 80 to 100% mycelial growth inhibition in our study (Fig. 3A). Similar result was reported that benomyl was not a highly effective preventative spray for melanoses (Whiteside, 1977). Work on post-infection mechanism of fungicides revealed that benomyl efficiently suppresses the pycnidiospores production on dead twigs rather than having less protectant activity (Bushong and Timmer, 2000; Whiteside, 1977). Disease resistance to fungicides may be now widespread. It is a disadvantage with the systemic products that act on one site only of the fungus (Finch et al., 2002).
Therefore, one resistant isolate NEL21-2 to benomyl may be in line with many studies of benomyl resistance (Altertini et al., 1999; Cooley and Caten, 1993; Koenraadt et al.,1992; Ma et al., 2003; Yarden and Katan, 1993). They reported that although benomyl resistance was common in fungi, resistance especially appeared after multiple applications. Besides, point mutations in the β-tubulin gene that change the amino acid sequence at the Benzimidazole binding site were commonly associated with fungal resistance to benomyl (Koenraadt et al., 1992).
Prochloraz belongs to the demethylation inhibitors (DMI) fungicides. Its target is the heme iron of cytochrome P450 sterol 14α-demethylase (CYP51) and interferes with the ergosterol biosynthesis of the cell membrane of the pathogenic fungus. Ergosterol participates in an essential role in mediating the permeability, rigidity, and fluidity of fungal membranes (Becher and Wirsel, 2012). DMI fungicides may affect the function of fungal essential membrane-bound enzymes such as chitin synthase and some membrane-bound enzymes may be involved in the detoxification of reactive oxygen species (Becher and Wirsel, 2012). It was reported that P. digitatum conidia produced on PDA amended with prochloraz at 0.05 and 0.1 mg/l had much higher percentages of impaired cell membrane integrity, and mycelia grown on prochloraz-amended PDA had less ergosterol (Zhang et al., 2021). These results were consistent with the established modes of action for the DMI fungicides. However, in contradiction to the mode of action, prochloraz reduced rather than increased mycelial cell membrane permeability. The reason may be related to reduced mycelial growth. The present study demonstrated that 40 isolates of D. citri hyphae produced on PDA amended with higher concentrations (1.45 to 181.25 μg/ml) of prochloraz had hyphae growth inhibition with no growth or the slowest growth (Fig. 3E). Therefore, present results may be consistent with the above findings of DMI fungicides.
Although the mode of action has been not fully understood, fluazinam has been believed to be interrupting the fungal cell’s energy production by an uncoupling effect on oxidative phosphorylation (Brandt et al., 1992; Guo et al., 1991; Vitoratos, 2014). The results of a study provided a strong evidence that the mode of action of fluazinam consists of the inhibition of the fungal cell’s energy production process through direct inhibition of the ATP synthetase (Vitoratos, 2014). Resistance development of fungicides with a specific mode of action may be a major threat to effective chemical control. This treatment may as well concern fluazinam because chemical resistance to the fungicide has only been described in field isolates of Botrytis cinerea (Tamura, 2000). However, no data related to its field resistance or the pattern of the cross-resistance of fluazinam with other respiration inhibitors have been reported so far (Lemay et al., 2002; Morita and Tojo, 2007). Our study also showed the in-vitro effectiveness of fluazinum on the hyphal growth of 40 D. citri isolates and there have no resistant isolates (Fig. 3C). Bushong and Timmer (2000) also reported that protectant fungicides such as fluazinam were more effective in controlling citrus melanose than curative fungicides. In fungicide resistance monitoring of jujube anthracnose, it was observed that fluazinam had an excellent inhibitory effect on mycelial growth and spore germination among the six tested fungicides. The EC50 value of fluazinum was 0.02 μg/ml, which was effective even at very low concentrations in that study (Lee et al., 2019). Similarly, in our study, ED50 of fluazinum was 0.18 μg/ml for 40 D. citri isolates (Fig. 3D).
Fungicide resistance monitoring of 40 representative D. citri isolates to kresoxim-methyl, benomyl, fluazinam, and prochloraz manganese with seven different concentrations showed 12 isolates as resistant to kresoxim-methyl, which could not inhibit the mycelium growth to more than 50% in vitro test, and also one isolate as resistant to benomyl, whose hyphae grew well even on the highest chemical concentration and inhibition was 0.98%. However, fluazinam and prochloraz manganese with no chemical resistance in fungal isolates were found. On the other hand, in vivo bio-testing efficacy on young citrus leaves of some resistant isolates was performed against both kresoxim-methyl and benomyl.
In conclusion, kresoxim-methyl showed less effectiveness on hyphal growth of D. citri and 12 isolates were resistant to kresoxim-methyl, which could not inhibit the mycelium growth to more than 50% in vitro. And one isolate was resistant to benomyl, whose hyphae grew well even on the highest chemical concentration. However, fluazinam and prochloraz manganese with no chemical resistance in fungal isolates were found. On the other hand, in vivo bio-testing on young citrus leaves of some resistant isolates was performed against both kresoxim-methyl and benomyl. These results showed that there are some resistant isolates of D. citri against both systemic fungicides, which can help screening out the commercial fungicides in the citrus production area.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement1
Supplement1 Print
Print






