



Apples, which grow in temperate regions, are one of the most important fruits worldwide, both economically and culturally. A total of 62 pathogens capable of infecting the fruit, trunks, and roots of apple trees, have been reported in Korea so far (Korean Society of Plant Pathology, 2023). To prevent economic losses, the Korean apple fungicide spray program was developed by prof. Emeritus Jae-Youl Uhm (Lee et al., 2009). However, due to climate change and newly emerged pathogens, it is difficult to maintain tree vigor and produce high-quality fruit (Das et al., 2020; Kim et al., 2018; Lee et al., 2017, 2021a, 2021b; Park et al., 2018). Recently, it has been shown that apple trees can be affected by unexpected decline, a worldwide phenomenon referred to as rapid apple decline (RAD). It was firstly reported in the United States, but the specific causative agent responsible for it has not been identified yet (Peter, 2018; Rosenberger, 2018; Singh et al., 2019; Stokstad, 2019). Reports have shown that RAD occurs mainly in young trees (2-8 years old) on Malling 9 (M.9) dwarf rootstocks and is characterized by various symptoms, such as stunted growth, pale yellow to reddish leaves, rolled and spindle leaves, peeling bark, necrosis around and above the rootstock-scion graft union, canker, and eventually decline within a weeks after the onset of symptoms (MacDonald et al., 2019; Peter, 2018; Rosenberger, 2018). Another common characteristic observed in affected orchards is the lack of a specific pattern in the distribution of infected trees, which appear to be evenly scattered among the healthy ones across the orchard (Peter, 2018; Singh et al., 2019). Several factors have been associated with RAD, and it is believed that multiple causes may contribute to its occurrence (Rosenberger, 2018; Singh et al., 2019).
Recently, RAD has been observed in apple orchards in Korea regardless of cultivar, cultivation period, or region. Such a widespread occurrence has led to misdiagnosis and confusion among farmers in regard to its causes. Several studies have been conducted to determine the factors that can be associated with RAD or apple decline more generally (Avalos-Ruiz et al., 2022b; Lee et al., 2016, 2020b, 2021a, 2023). However, due to the complex nature of RAD, it is difficult to establish the causative agents and what triggers this phenomenon. This review aims to summarize the current research into apple decline, including RAD worldwide, and suggest potential causal factors. In addition, the results of a survey conducted in apple orchards in Korea were presented in order to gain a further understanding of RAD.
Current Status of RAD Worldwide and Contributing Factors
RAD has been recently observed and studied in the United States and Canada, and it has similarities with a phenomenon known as quick decline (QD) in Japan (Fujikawa et al., 2019; Peter, 2018; Rosenberger, 2018). Currently, several factors, both biotic and abiotic, have been proposed to cause RAD worldwide (Table 1). Most studies have isolated opportunistic fungal pathogens from trees affected by stem canker, dieback, or decline. However, apple decline cannot be explained by a single factor (such as infection by fungal pathogens), and more studies are being conducted into more complex factors related to this phenomenon, such as fungal, bacterial, and viral infections (biotic factors) and extreme cold, insufficient soil moisture, drought, injuries caused by insects or rodents, and water stress (abiotic factors).
Abiotic factors
Environmental stressors, such as extreme temperature fluctuations, drought, or excessive moisture, may play a significant role in RAD development. These stressors can weaken apple trees, making them more susceptible to diseases. The occurrence of RAD could be attributed to variations in abiotic factors, including moisture levels in deteriorating roots, coupled with extreme weather conditions (Singh et al., 2019). In regard to drought stress, apple trees that experience drought in summer are susceptible to the opportunistic fungus Botryosphaeria dothidea, which resides in older bark and can potentially infect both the bark and cambium of water-stressed trees (Rosenberger, 2018). Moreover, trunk damage can manifest on the southwest-facing side of trees during winter due to heating-cooling cycles. Specifically, this occurs when solar radiation, sometimes amplified by reflection on the snow cover, heats the trunks on the southwest-facing sides during the day, and rapid cooling to sub-freezing temperatures follows at night. Factors such as excessive nitrogen fertilizer and drought conditions can make young trees more susceptible to winter damage by stimulating late-season growth (Rosenberger, 2018). Exposure to freezing conditions can lead to frost injuries, weakening trees and making them susceptible to pathogen infection and canker (Kennelly et al., 2007; Sobiczewski and Jones, 1992; Weaver, 1978). Overall, environmental stressors, like extreme temperature fluctuations or drought, can significantly contribute to RAD development in apple trees. Weakened trees become more vulnerable to diseases and pests, necessitating proper management to maintain tree health and productivity.
Biotic Factors
Bacteria
The plant pathogenic bacteria Dickeya dadantii and Pseudomonas syringae pv. syringae have been reported as the causative agents of apple QD and stem canker (Fujikawa et al., 2019; Lee et al., 2023). Specifically, QD was associated with infection by D. dadantii (formerly known as Erwinia chrysanthemi), which was isolated from the necrotic tissues and exudates of apple trees growing in some areas of Iwate Prefecture, Japan, and showing acute defoliation, red exudates from the trunk near the ground, and necrosis of leaves and branches (Fujikawa et al., 2019). In contrast, P. syringae pv. syringae (PSS) was isolated from apple trees affected by canker on the stems in various orchards located in Gyeongbuk province (Yeongju, Yeongyang, Cheongsong, Mungyeong, Andong, and Yechone), Korea (Lee et al., 2023). Interestingly, this pathogen not only causes disease in apple trees but also shows ice nucleation activity (INA), which can induce cold injury (Lee et al., 2023; Lim et al., 2019b).
Viruses
The common latent viruses in apple trees are the Apple stem pitting virus, Apple stem grooving virus, Apple chlorotic leaf spot virus, and Apple mosaic virus (Lee et al., 2020a; Rosenberger, 2018). However, these virus types are generally not directly associated to apple decline (Rosenberger, 2018). Recently, in addition to them, a new virus, known as Apple luteovirus 1 (ALV-1), has been identified in declined trees in the United States (Liu et al., 2018) as well as in Korea, Belgium, and Greece (Fontdevila Pareta et al., 2022; Lim et al., 2019a; Malandraki et al., 2020). Luteoviruses target only the phloem tissue, affecting sieve elements, companion cells, and the phloem parenchyma (Ali et al., 2020). The effects produced on these components are suspected to contribute to apple decline, however, further studies are needed to confirm this hypothesis. Recently, twenty previously described plant viruses and one viroid, as well as a new ilarvirus named Apple ilarvirus 2 (AIV2), have been identified from apple orchards affected by rapid decline in the Okanagan and Similkameen Valleys of British Columbia, Canada (Xiao et al., 2022). Among these viruses, none has been definitively confirmed as the primary causative agent of RAD. Therefore, it is important to conduct further extensive research to clarify the etiology of this disease.
Fungi
Numerous pathogens have been reported to cause stem canker, dieback, or decline in apple trees in several countries worldwide (Table 1). The frequently isolated fungal pathogens that may be associated with RAD belong mostly to genera Botryosphaeria, Diaporthe, Diplodia, Cytospora, and Neofusicoccum. Various infections have been reported in apple trees, however, the responsible fungi were known to be typical opportunistic pathogens. For this reason, while it is simple to isolate fungal pathogens from declined trees, it remains difficult to conclude that they cannot be major contributors to RAD, except for soil-borne fungal pathogens like Helicobasidium mompa, Phytophthora spp., Pythium spp., Rosellinia necatrix, or Athelia rolfsii. Recently, Fusarium tricinctum has been reported as the causal agent of apple decline in China (Zhang et al., 2022). In addition, studies have shown that the ice nucleation-active F. tricinctum and Fusarium diversisporum isolated from declined apple trees in Korea may be linked to RAD (Avalos-Ruiz et al., 2022a, 2022b). Thus, further research is needed for a comprehensive understanding of RAD as well as the identification of causal agents, which will contribute to combating this disease worldwide.
Factors Linked to RAD: Cold Stress Accelerated by Ice Nucleation-Active Microorganisms
Ice nucleation is the process by which ice crystals form, and INA refers to the capability of external particles to induce the freezing of water at temperatures higher than approximately −38°C, the freezing point of ultra-pure water (Failor et al., 2021; Kunert et al., 2019). Certain biological particles, including bacteria and fungi, have demonstrated their significance as ice nuclei, facilitating ice formation at temperatures ranging from −15°C to 0°C (Kunert et al., 2019; Lagzian et al., 2014; Pouleur et al., 1992). However, INA can play a role in ice formation within plants, potentially influencing the severity of frost damage. Previous studies have indicated that Pseudomonas syringae showed INA, which can cause frost damage to susceptible plants (Lindow et al., 1978). In Korea, PSS WSP007 was isolated from apple stem canker, and draft genomic analysis shed light on its life cycle, revealing that it exhibited INA (Lee et al., 2023; Lim et al., 2019b). At the same time, Fusarium strains obtained from declining apple trees in Korea have also been reported to show INA (Avalos-Ruiz et al., 2022a, 2022b). However, our understanding of the specific effects of INA caused by PSS or Fusarium species on different apple rootstocks depending on climatic conditions and geographic regions is still limited. Further research is necessary to increase our knowledge of the prevalence and significance of INA in apple orchards and to explore potential strategies for mitigating the detrimental effects of RAD.
Status of RAD in Korea
During the past decades, apple decline has been continuously observed in apple orchards in Korea. A variety of symptoms have been reported, including wilted flower and leaves, poor growth with small, spindle and curled leaves, yellowing, necrosis of the grafted union, and eventual decline regardless of cultivar or cultivation period (Fig. 1). Until now several fungal pathogens have been shown to affect the vigor of apple trees in Korea, including Botryosphaeria spp., Phytophthora spp., Cytospora mali (= Valsa mali), Diaporthe eres (= Phomopsis mali), Rosellinia necatrix, and Helicobasidium mompa, among others (Korean Society of Plant Pathology, 2023). Previous studies focused on isolating and identifying pathogens from declined apple trees. For instance, Valsa ceratosperma, P. mali, B. dothidea, and R. necatrix have been identified from diseased apple trees during a survey conducted from 1992 to 2000 in seven regions of Korea (Lee et al., 2006). Other studies isolated B. sinensis and B. kuwatsukai from apple trees experiencing decline (Lee et al., 2021a; Lim et al., 2023). Based on research conducted in various provinces of Korea, the most prevalent causal agents associated with apple decline symptoms were Phytophthora rot and violet root rot. Rodents, white root rot, and freezing injury were also found to be associated with apple dieback in 27 orchards between 2013 and 2019 (Lee et al., 2016, 2020b).
Between 2020 and 2022, we surveyed RAD-associated fungal pathogens on 91 declined apple trees from 48 orchards across seven Korean provinces. The most predominant infections observed were Botryosphaeria canker (63%) caused by Botrosphaeria spp., followed by Phomopsis canker (20%) caused by Diaporthe eres, Cytospora canker (9%) caused by Cytospora mali, white root rot (6%) caused by Rosellinia necatrix, and violet root rot (4%) caused by Helicobasidium mompa (data not shown). The findings highlighted the frequent presence of pathogenic fungi, especially Botryosphaeria spp., that can be associated with apple decline, which was in line Rosenberger (2018). Moreover, samples with a suspected Phytophthora root rot infection were assessed using the Agdia ImmunoStrip kit for Phytophthora, and 8 out of 27 of them resulted positive to the disease (data not shown). Phytophthora spp. are considered as potentially associated with apple decline but cannot be the sole factor explaining RAD.
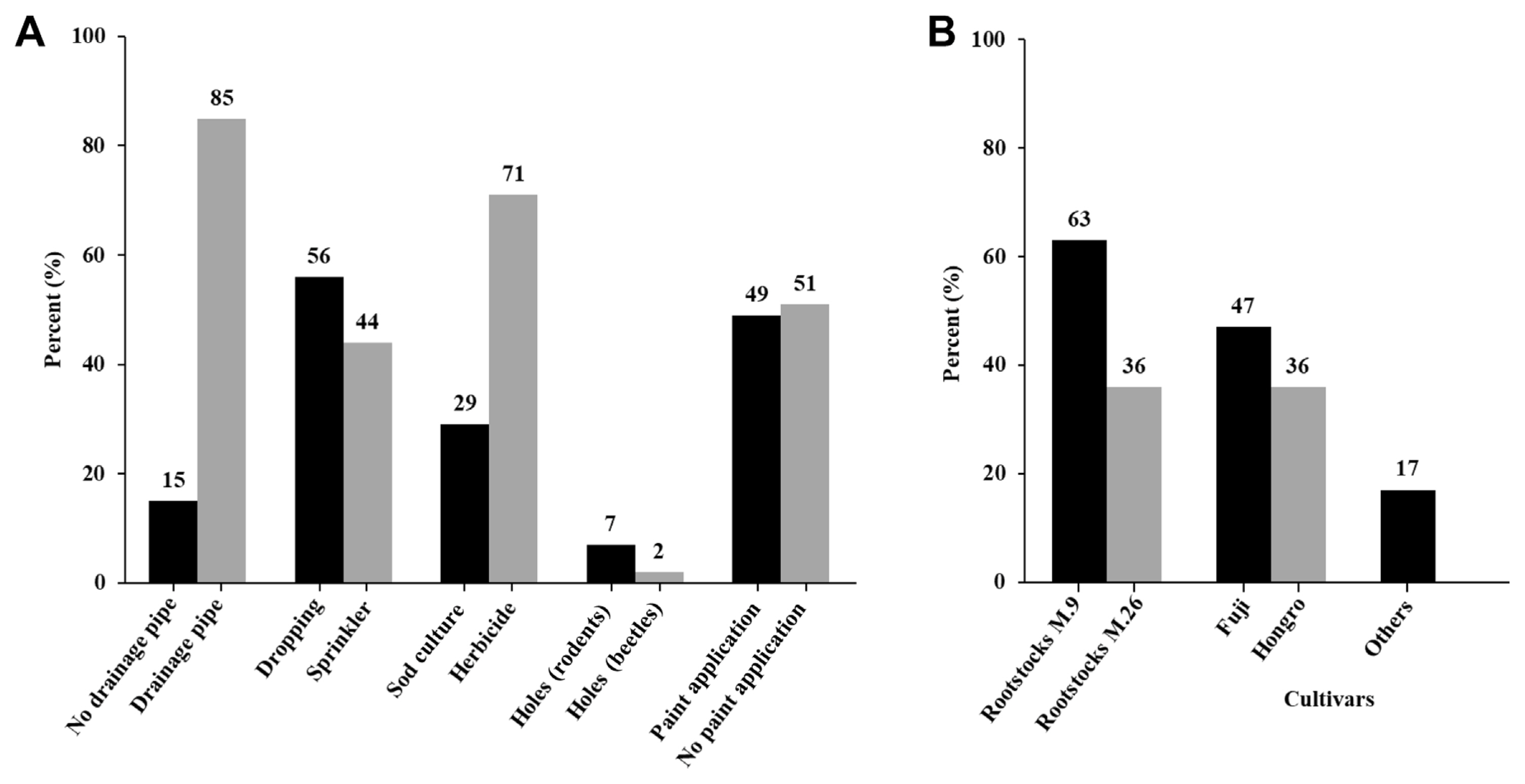
Cultivation techniques are important in the cultivation of apple trees. To examine the conditions of apple orchards affected by RAD, we assessed cultivation methods in 112 apple orchards between 2021 and 2022. Specifically, we examined the presence of drainage pipes under the orchards, type of irrigation, presence of beetles, application of white paint on trees, weed control methods, and kinds of rootstocks and apple cultivars (Fig. 2). The results showed that in about 85% of the RAD-affected apple orchards drainage pipes were used, and in 71% of them herbicides were applied to control the weeds under the trees. Extreme cold followed by drought/water stress, or either abiotic factor acting individually, may not have directly caused RAD but may have debilitated the trees and facilitated the proliferation of insects and infection by opportunistic pathogens (Singh et al., 2019). As for the irrigation types, it was found that dropping irrigation and mini-sprinkler irrigation were used in 56% and 44% of apple orchards, respectively, during two consecutive years. Dropping irrigation was slightly more used than mini-sprinkler irrigation; however, these could be hardly considered as direct causes of RAD. The presence of beetles was observed in only 2% of the RAD-affected orchards, and the application of white paint on trees, which is known to prevent cold injuries, showed similar incidence (49% vs. 51%) of RAD. The choice of rootstocks was also found to be important, with the rates of M.9 and M.26 rootstocks being 63% and 36%, respectively, in the RAD-affected orchards. RAD was observed regardless of apple cultivar; however, more that 80% of trees belonging to cvs. ‘Fuji’ and ‘Hongro’ were infected, and these are the dominant cultivars used for apple cultivation in Korea. The results here reported suggested that rootstock vigor and tree water use strategies influenced scion stress responses. This implies the need for vigor-specific irrigation management to alleviate stresses and achieve production objectives for various rootstocks (Xu and Ediger, 2021).
Conclusions and Future Perspectives
RAD is a complex syndrome with unclear etiology that poses significant challenges to the apple industry, particularly affecting young apple trees grafted on dwarf rootstocks. While extensive research has shed some light on the factors contributing to RAD, there is still much to be elucidated in relation to its underlying causes, prevention, and management. RAD appears to be influenced by both biotic and abiotic factors, rootstock interactions, and environmental conditions (Fig. 3), whose interplay is likely to contribute to the manifestation and severity of RAD symptoms. It is evident that the combination of different factors can have synergistic effects, exacerbating the decline of apple trees.
Recently, the average winter temperature has been rising not only in Korea but also globally, and the frequency of cold waves has increased (Jeon and Cho, 2015; Laing and Binyamin, 2013). In Korea, dormancy breaking in apple trees occurs between January and early February (Kweon et al., 2013). After this period, cold injury is likely to affect the trees if they experience a cold wave. At present, the most commonly used rootstocks for apple cultivation are M.9 and M.26, and, in terms of their cold hardiness, M.9 rootstocks are known to be more vulnerable to cold injury than M.26 (Moran et al., 2011; Westwood and Bjornstad, 1981). Therefore, the above-mentioned factors may contribute to the susceptibility of M.9 to RAD and may explain the higher incidence of the disease observed in these rootstocks than in M.26 trees (Fig. 2). Furthermore, INA can be a factor of RAD, highlighting the potential role of ice nucleation-active microorganisms in the decline of apple trees. Research suggests that elevated INA levels may increase the susceptibility to cold injury (Avalos-Ruiz et al., 2022a, 2022b; Lee et al., 2023; Lim et al., 2019b), leading to rapid decline due to infection by opportunistic fungi. However, further research is needed to fully understand the complex interactions between INA and other contributing factors like opportunistic pathogens.
In conclusion, RAD remains a complex syndrome requiring further research efforts to unravel its underlying causes, develop effective management strategies, and mitigate its impact on the apple industry. Collaboration between researchers, growers, and industry stakeholders is essential to address these knowledge gaps and safeguard the sustainability of apple orchards in the face of RAD.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






