

Introduction
Yield loss and low quality products of watermelon were reported in Korea in 2015, and the cause was usually the seed-transmitted virus Cucumber green mottle mosaic virus (CGMMV, genus Tobamovirus, family Virgaviridae), and which is a major viral cause of yield loss in cucurbits. CGMMV is not only the major seed-transmitted virus in watermelon (Citrullus lanatus) but also in cucumber (Cucumis sativus) and squash (Cucurbita moschata) (Lovisolo, 1979). CGMMV transmission is mainly by direct mechanical contact through contaminated soil, propagation stock materials and seeds (Choi, 2001; Hollings et al., 1975; Wu et al., 2011).
Many plant viruses are known to be seed-transmitted, and many tobamovirus infections result from transmission through seed. Seed transmission occurs either inside the embryo, or by external contamination of the seed coat (non-embryonic seed transmission). Embryonic seed-transmitted viruses are able to infect the embryo directly from maternal tissue, or through infected pollen (Liu et al., 2014). Embryonic seed transmission infects the newly germinated seedling and most seed-transmitted viruses have been identified as embryo-infecting. Meanwhile non-embryonically-transmitted viruses are unable to invade the embryo, and are found on the surface of seeds or spread through the seed coat as a contaminant, but are able to be transmitted mechanically to the developing seedling. Viruses that undergo strictly non-embryonic seed transmission are carried on the surface of seeds or as contaminants in seed lots and are not detected in the embryo. In general, members of the genus Tobamovirus have been shown to be seed transmitted in a non-embryonic way (Agarwal and Sinclair, 1996; Hull, 2002), although CGMMV has been shown to also internalize to the seed (Reingold et al., 2015).
Another tobamovirus, Pepper mild mottle virus (PMMoV) is also reported to result in yield loss caused by seed transmission (Wetter and Conti, 1988). As with other members of the genus Tobamovirus, CGMMV and PMMoV have a single-stranded, positive-sense RNA genome encapsidated in rigid rod-shaped particles by a single type of coat protein (CP) (Adams et al., 2009). The genomic RNA contains a 5′ cap and a tRNA-like structure in the 3′ untranslated region, and encodes four polypeptides. The tobamovirus 128 kDa protein and read-through 183 kDa RNA-dependent RNA polymerase proteins translated from the genomic RNA are used for viral replication (Derrick et al., 1997), whereas the 30 kDa cell-to-cell movement protein (MP) and the 17 kDa CP are translated from 3′ co-terminal subgenomic RNAs (Takamatsu et al., 1987).
Many efforts have been applied for reduction or elimination of seed transmission. Either trisodium phosphate or dry heat can be used to reduce seed transmission of PMMoV and other tobamoviruses in Capsicum spp. The disadvantage of chemical treatment is that it is not sufficiently effective to sterilize the virus from infected seeds. Either type of treatment has the risk of reducing seed viability or longevity (Demski, 1981; Nagai, 1981; Rast and Stijger, 1987).
Therefore, new advances in seed processing are required to overcome these disadvantages in order to produce high-quality seeds. The effects of extracts of some plants as inhibitors of plant virus infection have long been known (Duggar and Armstrong, 1925; Jones et al., 1959; Kassanis and Kleczkowski, 1948; Verma et al., 1969), with at least some of the active factors, such as pokeweed antiviral protein, known to be ribosome-inactivating proteins (Endo et al., 1988). Several micro-organisms have also long been recognized to produce inhibitors of plant viruses, including Aerobacter aerogenes and Aspergillus niger (Johnson, 1938), and extracts of multiple fungal species tested by Gupta and Price (1950), including filtrates of Trichothecium roseum and Neurospora sitophila. At least two compounds were identified from T. roseum, of which one is a complex ester known as trichothecin, and another a polysaccharide containing a high proportion of galactose (Bawden and Freeman, 1952). Occasionally researchers have tried to apply biological treatments to reduce virus concentration on the seeds. Few methods have been reported to treat seed and protect plants from viruses, such as use of Acinetobacter strain KNF2022T isolated from tobacco plant roots, which showed inhibitory effects on Tobacco mosaic virus (TMV) (Lee et al., 2009), and Enterobacter asburiae, which induced resistance to Tomato yellow leaf curl virus (TYLCV) through plant growth promoting rhizobacteria (Li et al., 2016). Whereas seed treatment with some botanical extracts had some inhibitory effects on reducing tobamovirus concentration in mechanically-inoculated pepper and tomato seedlings, apparently resulting from induced resistance (Madhusudhan et al., 2011), no effects on seed transmission of the viruses was reported. However, previous reports lacked information on the mechanism(s) to inhibit virus using these biological control methods. In this research we investigated potential interactions between a soil-borne bacteria-produced product and viral proteins to control viral disease using the seed-transmitted tobamoviruses CGMMV and PMMoV.
Materials and Methods
Screening of antiviral bacteria
We screened for bacteria from the rhizosphere of tobacco (Nicotiana tabacum) on Mueller-Hinton agar (MHA; Difco, Detroit, MI, USA) media, at Daejeon, Republic of Korea. More than 1,000 colonies of bacteria were isolated based on colony appearance, and grown on MHA media for 24 h at 30°C for further analyses. The culture filtrate of isolates used for antiviral activity screening were prepared by growth in Mueller-Hinton broth (MHB; Difco) for 2 day at 30°C on a shaking incubator (180 rpm). The culture broth was centrifuged at 10,000g × for 30 min to remove bacterial cell and culture debris. N. tabacum cv. Xanthi-nc was used for local lesion assay of TMV infection in the greenhouse. For the virus inoculum, leaves (0.1 g) of N. tabacum cv. NC82, systemically infected with TMV, were ground in 20 ml of potassium phosphate buffer (0.02 M, pH 7.3). The sap was filtered through two layers of cheesecloth, and the extract then centrifuged at 3,000 rpm for 15 min to remove debris; the supernatant was used as TMV inoculum. Antiviral activity in the local lesion host was tested by using the half-leaf method (Choi and Lee, 1995; Fukami et al., 1981). The 20-fold dilutions of culture filtrates were applied with cotton swabs to the upper surface of one half leaf, while Mueller Hinton broth was applied on the remaining leaf half as control. The virus was inoculated 24 h after application of the culture filtrate by using ordinary carborundum (600 mesh) methods, with five half leaves inoculated per treatment. The symptoms on the inoculated plants was observed at 3 to 4 days after the inoculation, and the effects of the treatment were measured by comparing the number of local lesions on the treated and untreated (control) leaf halves. Antiviral activity was assayed using three tobacco plants per strain. The antiviral activity was described as mean ± SE. All data obtained from the repeated experiments in each test were expressed as means ± standard deviation. Differences were statistically evaluated using R package (3.3.2). One-way ANOVA followed by a Duncan’s multiple range test was applied for multiple comparisons.
Identification of the selected bacterial strain
The bacterial morphology was observed by scanning electron microscopy (HITACHI S4300N; Hitachi, Tokyo, Japan) and transmission electron microscopy (Philips CM20; Philips, Amsterdam, Netherlands). Cultural and physiological characteristics of the bacterial isolate were examined by the usual methods for its classification (Schaad et al., 2001). Carbon source assimilation was examined using the Biolog GN test kit (Biolog Inc., Hayward, CA, USA) according to manufacturer’s specifications. The genomic DNA for 16S rRNA gene sequencing and phylogenetic analyses were extracted and purified according to the method of Sambrook and Russell (2001). The 16S rRNA gene was amplified by PCR using two universal primers, 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) (Weisburg et al., 1991). The purified gene fragment was sequenced using a BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster City, CA, USA) and examined using an automated DNA analyzer (Applied Biosystems 3730xl DNA Analyzer; Applied Biosystems). The sequence obtained was compared with available 16S rRNA gene sequences from GenBank (http://www.ncbi.nlm.nih.gov/blast/) and EzTaxon (http://www.eztaxon.org/) (Kim et al., 2012). Phylogenetic analyses were performed using the MEGA6 (Tamura et al., 2013) with three algorithms, the neighbor-joining (Saitou and Nei, 1987), maximum-likelihood (Felsenstein, 1993) and maximum- parsimony (Fitch, 1971) methods. Bootstrap values were calculated based on 1,000 replicates (Felsenstein, 1985).
Comparison to type strain
The type strain of Pseudomonas oleovorans, ATCC 8062, was obtained from Korean Collection for Type Cultures (KCTC) in order to compare antiviral activity with the selected strain.
Origin and maintenance of virus strains
The virus strains of PMMoV and CGMMV used to test for anti-viral effects of bacterial culture extracts were collected from soils of fields used for growing pepper or watermelon in areas of Jeollanam-do province where these viruses were detected. PMMoV and CGMMV were propagated separately by mechanical transmission to N. benthamiana. In addition, in vitro transcripts of infectious clones of PMMoV (Han et al., 2017) and CGMMV (Park et al., 2017) were used to infect seedlings of pepper and watermelon, respectively, in order to obtain infected seeds for further testing.
Determination of seed transmission rate after treatment of pepper and watermelon seeds from infected plants with strain KBPF-004 culture supernatants
To compare seed transmission in P. oleovorans strain KBPF-004 supernatant-treated and non-treated PMMoV-and CGMMV-infected seeds, we used virus infected seeds harvested from infected pepper and watermelon, respectively. To obtain virus infected seed, infectious clones of PMMoV (Han et al., 2017; J-76 KX399389) and CGMMV (Park et al., 2017; OMpj KX443592) were used. In vitro transcripts of each infectious clone were inoculated to seedlings of pepper and watermelon respectively. Healthy plants, and inoculated plants which were confirmed as infected by RT-PCR at 10 dpi, were transplanted in greenhouse. After 60 days pepper and watermelon seeds were harvested, cleaned, and dried at room temperature for 48 h. Dried seeds were then soaked in supernatant of strain KBPF-004 or control ATCC 8062 (each at OD600: 0.2) in 50 ml Falcon tubes with shaking for 20 min and dried again for 48 h at room temperature. Control seed were washed in the same manner, with water instead of culture supernatant, and similarly dried at room temperature. More than 300 dried seeds per treatment were directly planted in pots containing garden grow soil to germinate, in two separate experiments. At 10 days after germination each whole plant was separated from soil, and total RNA was extracted for cDNA synthesis and RT-PCR essentially as described (Han et al., 2017; Park et al., 2017) for PMMoV detection using primers PMMoVCP-F (5′-AAGGTGTTGATACCCAAGTTT-3′) and PMMoV-CP-R (5′-CAACGAGTTCTCGACTGAC-3′), yielding a product of 498 bp; or CGMMV detection using primers CGMMV-CP-F (5′-TTATGACATCGGCGGTAACT-3′) and CGMMV-CP-R (5′-GATTGTAAGCCATCTTCAAA-3′), yielding a product of 456 bp.
Subcellular localization of MP, CP and 126 kDa Protein of PMMoV and CGMMV after treatment with strain KBPF-004 culture supernatants
Green fluorescent protein (GFP) fusion proteins of the MP, CP and 126 kDa protein of PMMoV and CGMMV in binary vector pGDG (Goodin et al., 2002) were prepared as reported (Han et al., 2017; Park et al., 2017). The GFP:126 kDa, GFP:MP and GFP:CP of PMMoV, and GFP:CP of CGMMV binary vector constructs were grown in A. tumefaciens GV2260, diluted to OD600: 0.6, and infiltrated to N. benthamiana (Bragg and Jackson, 2004) in combination with an equal volume of various ratios of culture supernatants of strain KBPF-004 and ATCC 8062 at equivalent concentration. N. benthamiana leaves were divided into regions infiltrated with A. tumefaciens expressing individual GFP fusion constructs mixed with either strain KBPF-004 or ATCC 8062, essentially as shown in Li et al. (2014). At 2 dpi pieces of N. benthamiana leaves were collected for fluorescence detection. Confocal microscopy using a Zeiss LSM 710 (Zeiss, Oberkochen, Germany) detected green fluorescence with excitation at 488 nm from an argon laser; emission was monitored at 500-520 nm.
Western blot detection of GFP fusion proteins in agroinfiltrated leaves
Total proteins were extracted from agro-infiltrated leaves of N. benthamiana, separated by electrophoresis, blotted to PVDF membranes, and GFP fusion proteins detected using anti-GFP Living Colors monoclonal antibody (Clontech, Mountain View, CA, USA), as previously reported (Han et al., 2017).
Results
Screening and identification of antiviral bacteria
First we isolated more than one thousand colonies of soil-borne bacteria collected in or near tobacco fields located in Daejeon, Republic of Korea. The isolated soil bacteria were grown in MHB media, and supernatants of bacterial cultures were used for antiviral activity screening. Through this antiviral assay we selected nine bacterial isolates as shown in Table 1. Among these nine selected soil-borne bacteria, isolate KBPF-004 showed the highest antiviral activity when 1/20 diluted supernatant was used to treat tobacco leaves, with a mean reduction of 85.5% in local lesion numbers compared to the control non-treated leaf regions (half-leaf method) (Table 1). The remainder of the soil-borne bacteria isolates did not show any significant antiviral activity. Isolated strain KBPF-004 was identified and assigned development code KNF2016.
Identification of bacterial strain
Isolated strain KBPF-004 was an aerobic, motile, Gram-negative organism. The young colonies had circular form, convex with entire margins, moist, shiny and yellow-color. Older colonies became rough and uplifted, star-shaped and with dark yellow color. Cells were rods of about 0.5-1.0 μm in width by 1.5-3.0 μm in length at 24 h incubation on MHA, and coccoid cells occurred in old cultures. The temperature range for growth was 10-37°C with optimal growth at 30°C. The pH range for growth was pH 6.0-8.0 with optimal growth at pH 7.0. No growth occurred with an NaCl concentration of more than 5%, whereas optimal growth occurred without added NaCl. The major fatty acids in the isolate were 18:1 ω7c, 16:0, and summed feature 3, which contained one or more of the following fatty acids: 16:1 ω7c and/or 15:0 iso 2OH. The major quinone of strain KBPF-004 was Q-9. The 16S rRNA (1,473 bp) sequence of strain KBPF-004 showed 99.05% identity to that of P. oleovorans strain IAM 1508T (accession no. CP000560). The strain KBPF-004 was deposited at the KCTC as P. oleovorans strain KBPF-004 (KCTC 10159BP). Similar analysis of the type strain of P. oleovorans, ATCC 8062 was performed in order to compare with strain KBPF-004; ATCC 8062 showed 19.7% antiviral activity, whereas strain KBPF-004 showed 96.0% (Table 2). This indicated that strain KBPF-004 has antiviral characteristics significantly different from those of the P. oleovorans type strain.
Infectivity and seed transmission rate of PMMoV- and CGMMV-infected seeds following treatment with supernatant of P. oleovorans strain KBPF-004 cultures
We have previously detected seed-transmission of up to 70% among germinated seeds with CGMMV (Park et al., 2017) and similar levels with PMMoV (Han et al., 2017). In vitro transcripts of PMMoV and CGMMV infectious clones were inoculated to seedlings of pepper and watermelon respectively. Harvested seeds were soaked in culture supernatants of either strain KBPF-004 or control strain ACTC 8062 and about 300 seeds of each treatment planted. After twenty days, RNA was extracted from germinated seedlings to detect viral RNA by RT-PCR (Table 3). Significantly fewer germinated seeds treated with P. oleovorans strain KBPF-004 were infected with PMMoV or CGMMV compared to seed treated with strain ATCC 8062. The seed transmission rate of PMMoV was decreased from 61.9% (ATCC 8062 treated) to 15.5% (KBPF-004 treated) and of CGMMV from 59.7% to 15.8% (Table 3), indicating that products of P. oleovorans strain KBPF-004 directly affected viability of CGMMV and PMMoV. Infection rates of ATCC 8062-treated seed were not significantly different from the water wash control (Table 3).
Effects of supernatant of P. oleovorans strain KBPF-004 on 126 kDa, MP and CP of PMMoV
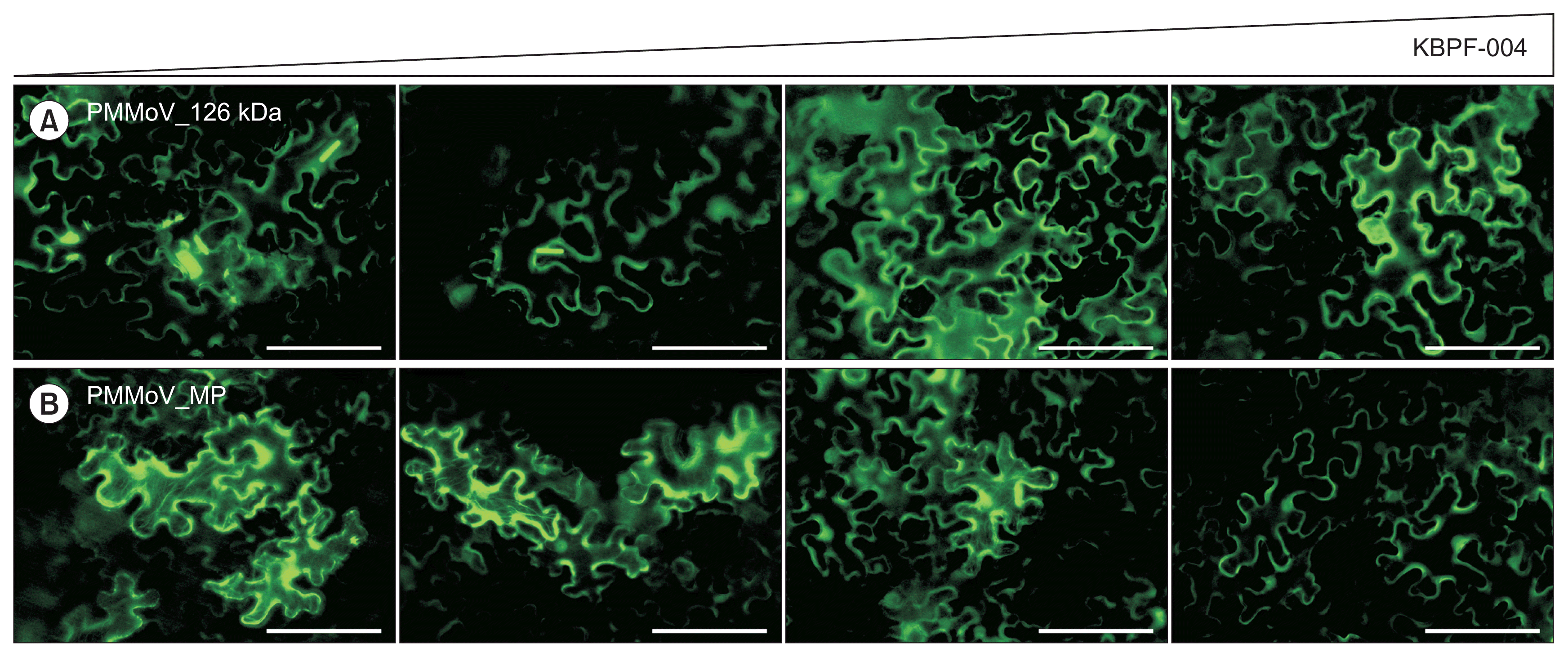
Previously we have reported the subcellular localization of agroinfiltrated GFP fusions of the 126 kDa protein, MP, and CP of isolates of CGMMV (Park et al., 2017) and PMMoV (Han et al., 2017). In contrast to results reported for other tobamoviruses (Liu et al., 2005) the PMMoV isolate used in this report showed a different localization pattern of 126 kDa ORF1 (Han et al., 2017). Except for the 126 kDa protein, localization of all GFP fusion proteins followed similar patterns to those reported for other tobamoviruses. When GFP:126 kDaPMMoV, GFP:MPPMMoV, and GFP:CPPMMoV were expressed with different ratios of KBPF-004 and ATCC 8062 supernatants, some differences were observed. In contrast to the normal localization pattern of GFP:126 kDa showing strong aggregation in the cytoplasm in the presence of ATCC 8062 supernatant, we observed that the aggregation diminished in N. benthamiana leaf agroinfitrated with increasing proportion of strain KBPF-004 supernatant (Fig. 1A, 2A). As with GFP:126 kDa, GFP:MPPMMoV localization at the cytoskeleton became weaker at higher proportions of strain KBPF-004 supernatant (Fig. 1B, 2B). Unlike the 126 kDa and MP proteins, the localization pattern of the GFP:CP did not change (Fig. 1C), but GFP:CP detection was significantly reduced in the western blot in P. oleovorans strain KBPF-004-treated compared to the ATCC 8062-treated control (Fig. 1E). Western blots showed that levels of the ORF1 and MP proteins did not appear to differ between treatments (data not shown). Interestingly, the pattern of localization of GFP:CPPMMoV did not show any difference, but the western blot suggested that the amount of CP decreased in the presence of P. oleovorans strain KBPF-004 supernatant, and similar results were observed for GFP:CPCGMMV (Fig. 1D, 1E). The subcellular aggregation or localization patterns of 126 kDa and MP of PMMoV were affected more in the presence of an increased proportion of supernatant of P. oleovorans strain KBPF-004 (Fig. 2). A reduction in the size of GFP:126 kDa aggregates was observed at higher proportions of KBPF-004 supernatant, with increased distribution at the cell periphery (Fig. 2A). For GFP:MP, as the proportion of KBPF-004 supernatant increased, there was reduced association between the MP and the cytoskeleton (Fig. 2B; left to right).
Discussion
Occasional reports of soil-borne bacteria having antiviral functions exist, such as that of Acinetobacter antiviralis against TMV (Lee et al., 2009); however, the detailed mechanism was not clearly understood. In this study we have focused on antiviral activity of candidate bacteria collected from field soils contaminated with tobamoviruses. P. oleovorans has been reported as an oil degrading bacterium (Lee and Chandler, 1941), but here for the first time we have demonstrated that isolate strain KBPF-004 (developmental code KNF2016) has significant antiviral activity not present in type isolate ATCC 8062. Firstly, we were able to screen antiviral activity of a soil-borne bacteria against TMV using a half-leaf assay. Secondly, through our experiments with tobamovirus-infected seeds, we observed significantly reduced seed transmission rates of both PMMoV and CGMMV following seed treatment with P. oleovorans strain KBPF-004 supernatant. Our further experiments to examine expression levels and subcellular localization of individual gene product:GFP fusion proteins in the presence of an increasing proportion of P. oleovorans strain KBPF-004 supernatant contribute to an understanding of these results. Tobamoviruses are often highly seed-transmitted, and the CGMMV and PMMoV isolates tested each showed about 70% seed transmission rates (Han et al., 2017; Park et al., 2017) (Table 3). However, our data showed that transmission of these seed coat-contaminating viruses was significantly reduced by seed treatment with culture supernantant of P. oleovorans strain KBPF-004; as control strain ATCC 8062 did not show any significant reduction of seed transmissibility of CGMMV and PMMoV, we suggest that soluble bacterial products in the culture supernatant of KBPF-004 directly affect the viability of virions on the surface of watermelon and pepper seeds. In addition, expression of the PMMoV CP in plant cells through agroinfiltration in presence of diluted supernatant of P. oleovorans strain KBPF-004 showed a reduced amount of CP compared to ATCC 8062. The reduction in levels of CP detected after agroinfiltration of infectious clones of PMMoV or CGMMV in the presence of culture supernatant of KBPF-004, but not ATCC 8062, is interesting. This suggests several possible mechanisms of action for the antiviral effects of KBPF-004, including (a) proteolytic cleavage directed at the CPs of both PMMoV and CGMMV; (b) interference in the production of the subgenomic RNA (sgRNA) encoding the CP; (c) inhibition of CP sgRNA translation; or (d) interference in virion stability affecting either assembly or disassembly of viral particles leading to targeting of free CP for degradation by the proteasome. The inhibition of seed transmission after seed washing in culture supernatant could potentially also be explained by proteolytic cleavage of CP in virions, exposing the genomic RNA to ribonuclease digestion, although an alternate explanation could be surfactant activity releasing viral particles from the seed coat, with or without virion destabilization. Further experiments will be directed to differentiating between these possibilities, in order to elucidate the mechanism(s) of antiviral activity of KBPF-004 culture supernatant, and to determine whether the observed antiviral effects against the tobamoviruses PMMoV and CGMMV extend to viruses from other taxonomic groups.
In addition to the reduction of CP expression by ingredient(s) of the culture supernatant in agroinfiltrated leaves, subcellular aggregation of 126 kDa, and association of MP with the cytoskeleton were both diminished by co-infiltration with diluted supernatant. The altered or diminished functions of 126 kDa and MP caused by components in the culture supernatant suggest multiple mechanisms for the diminished infectivity of seed-borne PMMoV and CGMMV when treated seeds germinated.
More than ten thousand secondary products have been reported to have been dissolved in the bacterial growth media (Monod, 1949). Thus one or multiple ingredients in supernatant of P. oleovorans strain KBPF-004 could affect CP, MP, and 126 kDa proteins of CGMMV and PMMoV. Although we have not yet been able to identify the factors in P. oleovorans strain KBPF-004 culture supernatants responsible for the observed antiviral effects, we have established procedures by which we were able to (1) initiate a screen to identify antiviral microorganisms in soil from virus infected fields and (2) evaluate multiple specific antiviral activities through the expression of viral proteins in the presence of various proportions of antiviral candidate elements contained in culture supernatant of the microorganism examined in this report.
Further research will seek to identify the unknown product(s) from P. oleovorans strain KBPF-004 (KNF2016) which confer the observed antiviral activities, and to investigate both the mechanisms of action, and possible evolutionary interactions among antiviral microorganisms, virus infected plants, and virus-contaminated soil.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






