

Soybean [Glycine max (L.) Merrill], which is one of the most important crops worldwide, is an economically important legume used as a source of oil, feed, and protein (Hartman et al., 2011). Soybean production in 2016 exceeded 330 million tons (FAOSTAT, 2016). In Asian countries, including Japan, soybean is an essential crop for traditional food products (Soyinfo Center: www.soyinfocenter.com). Between 2007 and 2016, Japan annually produced 199.9-261.7 × 103 tons, which placed the country 17th-23rd in the global ranking of soybean producers (FAOSTAT, 2016). However, this production level accounted for only 21-25% of the yield needed for the self-sufficient production of soybean as food. Therefore, the soybean industry in Japan is largely dependent on soybean imported from countries such as the US, Brazil, and Canada (FAOSTAT, 2016). Nonetheless, there has been a continuous demand for domestically grown conventional soybean, but soybean production has not increased in Japan for several decades. The temporal variation in average annual prefectural yields between 1958 and 2016 ranged from 0.94 to 1.66 tons per hectare, and annual yields have not increased in 89% of the analyzed prefectures (Chen, 2018). The obstacles that have prevented improvements in soybean production have included diverse abiotic and biotic factors such as extremes in nutrient availability, temperature, and moisture as well as diseases, pests, and weeds (Hartman et al., 2011).
One of the biotic constraints that can decrease soybean seed yield and/or quality is Phytophthora root and stem rot caused by Phytophthora sojae Kaufmann and Gerdemann [synonyms: Phytophthora megasperma var. sojae, P. megasperma f. sp. glycinea, and P. sojae f. sp. glycines], which has threatened soybean production worldwide (Grau et al., 2004; Hansen and Maxwell, 1991; Kaufmann and Gerdemann, 1958; Schmitthenner and Dorrance, 2015; Tyler, 2007; Wrather et al., 2001a). In 1998, the soybean yield in the top-10 soybean-producing countries decreased by an estimated 1,266 × 103 tons because of Phytophthora root and stem rot (Wrather et al., 2001a). In the US, the second largest decrease in soybean yield (an estimated 3,709 × 103 tons) occurred between 1996 and 1998, and was caused by Phytophthora root and stem rot (Wrather et al., 2001b). A survey conducted from 1996 to 2007 revealed that Phytophthora root and stem rot was the second and third worst diseases in terms of yield losses in 7 and 4 out of the 12 years, respectively (Wrather and Koenning, 2009). In Japan, this disease spread to several prefectures within 10 years of its first occurrence in 1977 (Sugimoto et al., 2006, 2012), and has since become a nationwide concern among soybean producers. Significant yield losses attributed to the disease have been reported in the top-10 soybean-producing countries as well as in Australia and Japan (Schmitthenner and Dorrance, 2015).
Phytophthora sojae, which is a homothallic oomycete classified in the kingdom Stramenopiles (Dorrance, 2018; Förster et al., 1990; Harper et al., 2005; Tyler, 2007), grows saprophytically and readily produces sexual oospores in soil and infected plant tissue. The oospores can survive in soil and diseased plant debris for extended periods (Dorrance et al., 2007; Tyler, 2007), suggesting that soybean plants grown in fields previously infected with Phytophthora root and stem rot are continuously at risk of developing the disease. The mycelia grown in soil and infected plant tissue produce asexual spores called sporangia, in which motile zoospores are generated. The zoospores are released from the sporangia under waterlogged conditions, after which they differentiate to form adhesive cysts that can infect soybean plants. The pathogen repeats its life cycle via a secondary infection during a soybean growing season under favorable environmental conditions. Because the infection by zoospores is essential to initiate the parasitic phase of the P. sojae life cycle, fields saturated for a long time provide one of the most ideal conditions for inducing Phytophthora root and stem rot. In Japan, most soybean plants are cultivated in fields converted from rice paddies (Kato et al., 2013; Sugimoto et al., 2012). Such soybean fields usually contain poorly drained soil with a high moisture level, which is ideal for a P. sojae infection and subsequent multiplication. Kato et al. (2013) reported that P. sojae is involved in pre-emergence soybean seedling damping-off in fields converted from rice paddies. Thus, soybean cultivars that are susceptible to the disease require the application of cultural and/or chemical practices to control P. sojae.
Host resistance to P. sojae, which is one of the most economical and environmentally friendly disease control measures, has been well-studied, especially for major resistance (R) gene-mediated resistance involving the race-specific interactions between soybean and P. sojae (Dorrance, 2018; Dorrance et al., 2007; Grau et al., 2004; Sugimoto et al., 2012). Previous studies identified 20 or more R genes or alleles (Rps: resistance to P. sojae), some of which have been commercially deployed to manage the disease in soybean fields (Dorrance, 2018; Jiang et al., 2017). In addition to the Rps-mediated resistance, soybean plants express partial resistance (i.e., quantitative resistance, field resistance, rate-reducing resistance, or tolerance) in a race-non-specific manner that is effective against all P. sojae races (Anderson and Buzzell, 1982; Dorrance, 2018; Schmitthenner, 1985; Sugimoto et al., 2012; Tooley and Grau, 1982, 1984). Unlike Rps-mediated resistance, this incomplete resistance leads to different levels of susceptibility/resistance among cultivars and can be evaluated based on decreased infection or colonization efficiency, the development of smaller lesions, and the production of fewer oospores in infected plant tissue (Dorrance, 2018; Mideros et al., 2007). In the US, soybean germplasm exhibiting various levels of partial resistance has been identified by screening the plant introduction series of the soybean germplasm collection belonging to the US Department of Agriculture (Dorrance and Schmitthenner, 2000; Jia and Kurle, 2008; Schneider et al., 2016). Several quantitative trait loci have been detected (Abeysekara et al., 2016; Burnham et al., 2003; Lee et al., 2013a, 2013b; Schneider et al., 2016; Tucker et al., 2010; Wu et al., 2011). Meanwhile, partial resistance among Japanese soybean cultivars has been studied mainly in Hokkaido and Hyogo prefectures, and six cultivars and one cultivar, respectively, were selected as candidates exhibiting a high level of partial resistance (Sugimoto et al., 2012; Yamashita et al., 2012). However, the tested cultivars or lines were limited to those grown locally.
Partial resistance against Phytophthora root and stem rot is not expressed in the soybean seed, but is effective during the growth stage in which the first trifoliate leaf is formed as well as in the subsequent stages (Schmitthenner and Dorrance, 2015). The resulting primary symptom is decreased root colonization (Dorrance et al., 2007). Additionally, ‘Hark’ plants (i.e., susceptible cultivar) inoculated at 16 days after seeding are more resistant to Phytophthora root and stem rot than younger plants (Jimenez and Lockwood, 1980). Meanwhile, one of the best-studied cultivars regarding this type of resistance, ‘Conrad’, which lacks Rps genes, is initially susceptible, but then becomes highly tolerant to P. sojae (Fehr et al., 1989), with significantly less seedling emergence when plants are inoculated up to 5 days after seeding compared with inoculations at 7, 10, and 14 days (Dorrance and McClure, 2001). Thus, soybean plants without effective Rps genes are susceptible to P. sojae during the early vegetative growth phase when the characteristic disease symptoms, including seed rot and pre- and post-emergence damping-off of seedlings and young soybean plants, are induced by the pathogen. Accordingly, characterizing the interactions between soybean cultivars and P. sojae in the seedling and early growth stages is important for developing effective disease management strategies because the onset of disease in the early part of the soybean growing season leads to direct plant losses and the need for costly replanting to replace soybean stands.
In this study, we analyzed the disease responses of soybean cultivars currently grown in Japan, including the major cultivars in terms of planted area, during the early growth stages. Our investigation involved multiple P. sojae isolates from Japanese soybean fields. We also examined the efficacy of commercially available pesticides applied to seeds to provide protection against Phytophthora root and stem rot. The objectives of this study were to (i) reveal the differences in the resistance to Phytophthora root and stem rot among Japanese soybean cultivars during the early growth stages, and (ii) determine the effective period for seed treatments with pesticides. The data presented herein demonstrate that seed treatments involving effective pesticides are important for protecting soybean plants from a P. sojae infection during the seedling and early growth stages.
Materials and Methods
Phytophthora sojae cultures
The following five P. sojae isolates were obtained from symptomatic soybean plants collected in Ibaraki prefecture, Japan in 2006-2014: 13-B-RO-1, 13-B-RO-2, 13-7-B1-1, 13-8-11-2, and Ps1001 (Table 1). We isolated P. sojae as previously described by Sugimoto et al. (2006), but with some modifications to the contents of the V8 juice agar medium. Diseased soybean stems were washed with tap water or sterile distilled water, after which stem sections containing the edge of lesions and the adjacent healthy part were excised. The stem sections were immersed in 70% ethanol for 10-20 s and 0.5% sodium hypochlorite for 1-2 min and then rinsed twice with sterile distilled water. After air-drying on sterile filter paper, the stem sections were placed on BNPRA-V8 juice agar medium (200 ml V8 juice, 3 g CaCO3, 1.5% agar, 10 mg/l benomyl, 10 mg/l nystatin, 7.5 mg/l pentachloronitrobenzene, 5 mg/l rifampicin, and 500 mg/l ampicillin) supplemented with 50 mg/l 3-hydroxy-5-isoxazole. Samples were then maintained at 25°C in a Cool Incubator (Mitsubishi Electric Engineering Co., Ltd., Tokyo, Japan). Mycelial tips derived from the stem pieces were transferred to Petri plates containing fresh BNPRA-V8 juice agar medium supplemented with 50 mg/l 3-hydroxy-5-isoxazole. For each isolate, the elongating hyphal tips were transferred to fresh V8 juice agar medium and grown at 25°C in a Cool Incubator.
Four of the 17 isolates, Ps060726-2-1, Ps080806-3-5, Ps060626-4-1, and Ps060629-5-1, were collected from Ibaraki, Fukushima, Miyagi, and Toyama prefectures, respectively (Table 1), and were provided by the Hokuriku Research Center, Central Region Agricultural Research Center of the National Agriculture and Food Research Organization (CARC/NARO).
Eight P. sojae isolates were obtained from the soil collected from soybean fields in Akita, Nagano, Fukuoka, and Saga prefectures, Japan in 2014 (Table 1). The soil samples were maintained at room temperature in a plastic bag until used. ‘Nattosyoryu’ seeds were sown in plastic pots (1/10,000 a) filled with field soil. After 3-4 days under glasshouse conditions, the potted plants were flooded for 22 h. Two days later, plants with Phytophthora root and stem rot lesions were collected for a subsequent isolation of P. sojae according to the above-mentioned procedure.
A PCR analysis using a species-specific primer pair (PSOJF1 and PSOJR1) (Bienapfl et al., 2011) confirmed that the expected amplicon was produced for all isolates except for four isolates provided by the Hokuriku Research Center, CARC/NARO. The four isolates from the Hokuriku Research Center have been already identified as P. sojae by means of morphological observation, PCR-based molecular characterization, and inoculation of soybean seedlings (Moriwaki, 2010).
Plant materials and seed treatment with pesticides
Eighteen Japanese soybean cultivars were used in this study (Table 2). Nine of the 18 cultivars (‘Tachinagaha’, ‘Enrei’, ‘Fukuyutaka’, ‘Toyomusume’, ‘Ryuhou’, ‘Miyagishirome’, ‘Yukihomare’, ‘Satonohohoemi’, and ‘Osuzu’) were among the 10 most planted soybean cultivars in Japan in 2015 (i.e., in terms of cultivated area). The nine other cultivars were as follows: ‘Himeshirazu’, ‘Nattosyoryu’, ‘Suzumaru’, ‘Suzuyutaka’, ‘Tamahomare’, ‘Sachiyutaka’, ‘Toyokomachi’, ‘Tanbakuro’, and ‘Hatayutaka’. ‘Suzumaru’ and ‘Sachiyutaka’ were among the 10 most planted soybean cultivars in 2012 and 2012-2014, respectively. The statistical data was obtained from the governmental official website for Ministry of Agriculture, Forestry and Fisheries of Japan. The Rps genes have not been detected in these 18 Japanese cultivars, although some of the cultivars are reportedly resistant to certain P. sojae races (Jiang et al., 2017; Yamashita et al., 2012). Two US cultivars, ‘Williams’ and ‘Harosoy’, which lacks Rps genes (rps) and expresses low levels of partial resistance and possesses Rps7 (Anderson and Buzzell, 1992; Burnham et al., 2003; Wu et al., 2011), respectively, were also inoculated with Japanese P. sojae isolates.
Soybean seeds (approximately 25 seeds per cultivar) were sown in plastic pots (1/10,000 a) containing 700 ml vermiculite (Burnpiece, S size; Hakugen Earth, Tokyo, Japan), and covered with 200 ml vermiculite. The vermiculite in pots was completely moistened with water, after which the pots were placed on a plastic tray (Sanko vat No. 1, 7 cm deep; Sanko Co., Ltd., Gifu, Japan). Water was then supplied to the plants through a hole at the bottom of the pots starting from 2-3 days after seeding. The pots were incubated in a glasshouse at CARC/NARO as described by Kato et al. (2013) to prepare the soybean plants for the inoculation test. To investigate the effects of pesticides on P. sojae, ‘Nattosyoryu’ seeds were treated with three commercially available pesticides (Table 3) at the recommended rates. The plants resulting from the pesticide-treated seeds were prepared for the inoculation test according to the above-mentioned procedure.
Inoculation and evaluation
Soybean plants were inoculated with P. sojae cultured on V8 juice agar medium as described by Mukobata and Sekihara (2006), with some modifications (i.e., P. sojae-V8 culture flooding method). Specifically, seven small pieces (approximately 5 × 5 mm) of a P. sojae culture grown on V8 juice agar medium were used to inoculate fresh V8 juice agar medium in 9 cm plastic Petri plates (Ina-Optika Co., Ltd., Osaka, Japan). The plates were incubated for 14 days at 25°C in an incubator. The P. sojae-V8 culture was roughly cut into small pieces (approximately 2 × 2 cm) using a spatula and then added to the vermiculite in each pot. The pots were flooded to approximately 2 cm above the vermiculite surface and then incubated for 5 days in the glasshouse. The inoculated plants were grown for an additional 5 days, during which plants were watered through a hole at the bottom of the pots. The inoculated soybean plants were evaluated in terms of the number of dead plants per emerged plants. Example of the disease symptoms caused by the P. sojae inoculation is shown in Fig. 1. None of ‘Nattosyoryu’ plants treated with only V8 juice agar pieces died, while all ‘Nattosyoryu’ plants were killed by inoculation with the Ps060626-4-1-cultured V8 juice agar pieces (Fig. 1). The extent of the disease resistance among inoculation tests was compared based on the proportion of dead plants.
Data analysis
Data were analyzed using R software (version 3.4.4) (R Core Team, 2018). In the time course experiment, the rates of dead plants due to the P. sojae infection underwent an analysis of variance (ANOVA) for each soybean cultivar grown for a different number of days before being inoculated. To determine significant differences among the mean rates of dead plants, Tukey’s honestly significant difference (HSD) test was completed by inputting the ANOVA results into the glht function of an R package, multcomp. The rates of dead plants due to the P. sojae infection were also compared among the 20 soybean cultivars using the above-mentioned method. In the time course evaluation of the efficacy of pesticide treatments of ‘Nattosyoryu’ seeds, the rates of dead plants due to the P. sojae infection underwent an ANOVA for the non-treated control plants and the plants derived from seeds treated with one of three pesticides. Significant differences between the mean rates of dead plants for pesticide-treated and non-treated plants were determined by analyzing the ANOVA results with the glht function of multcomp according to Dunnett’s multiple comparison test.
Results
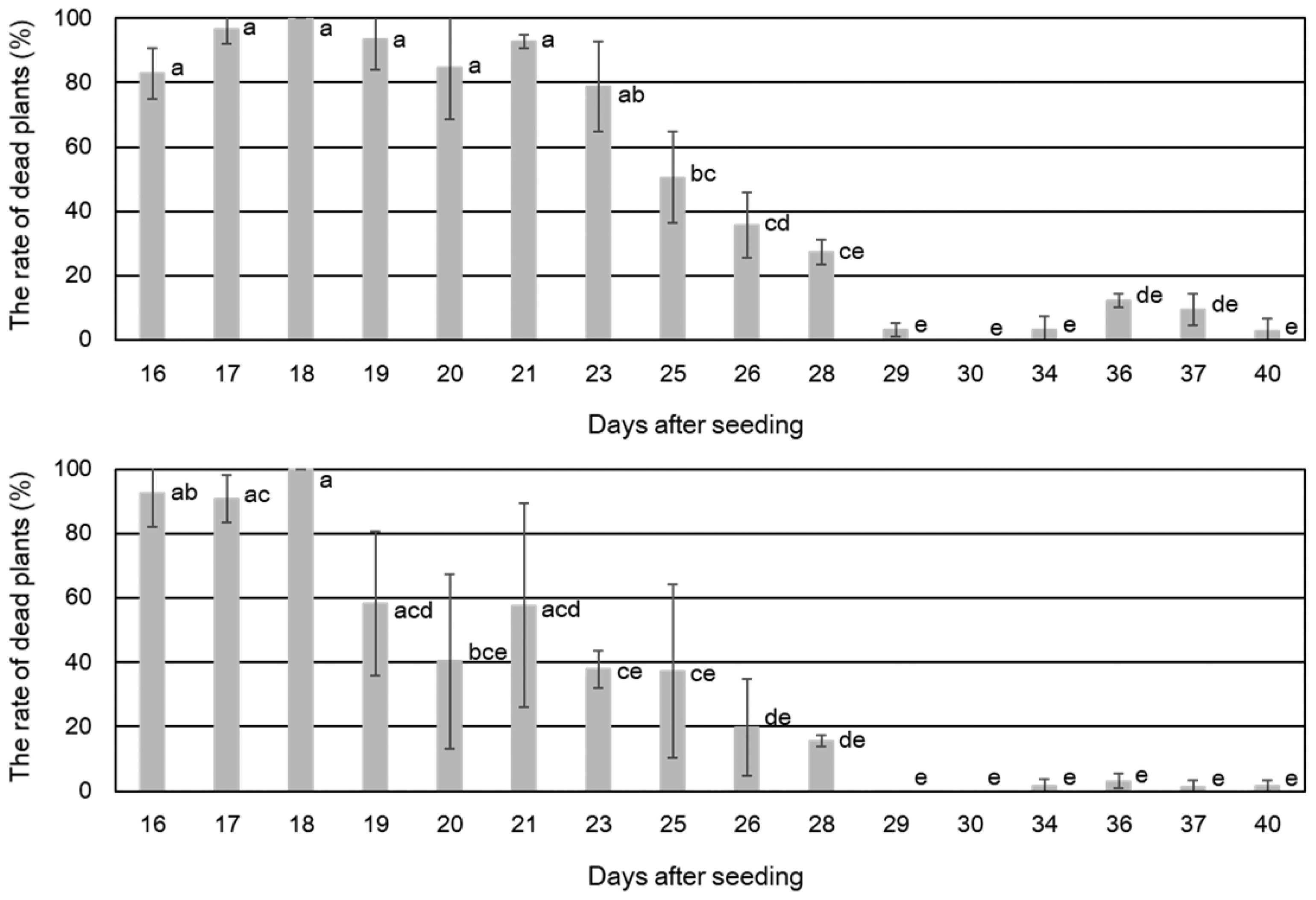
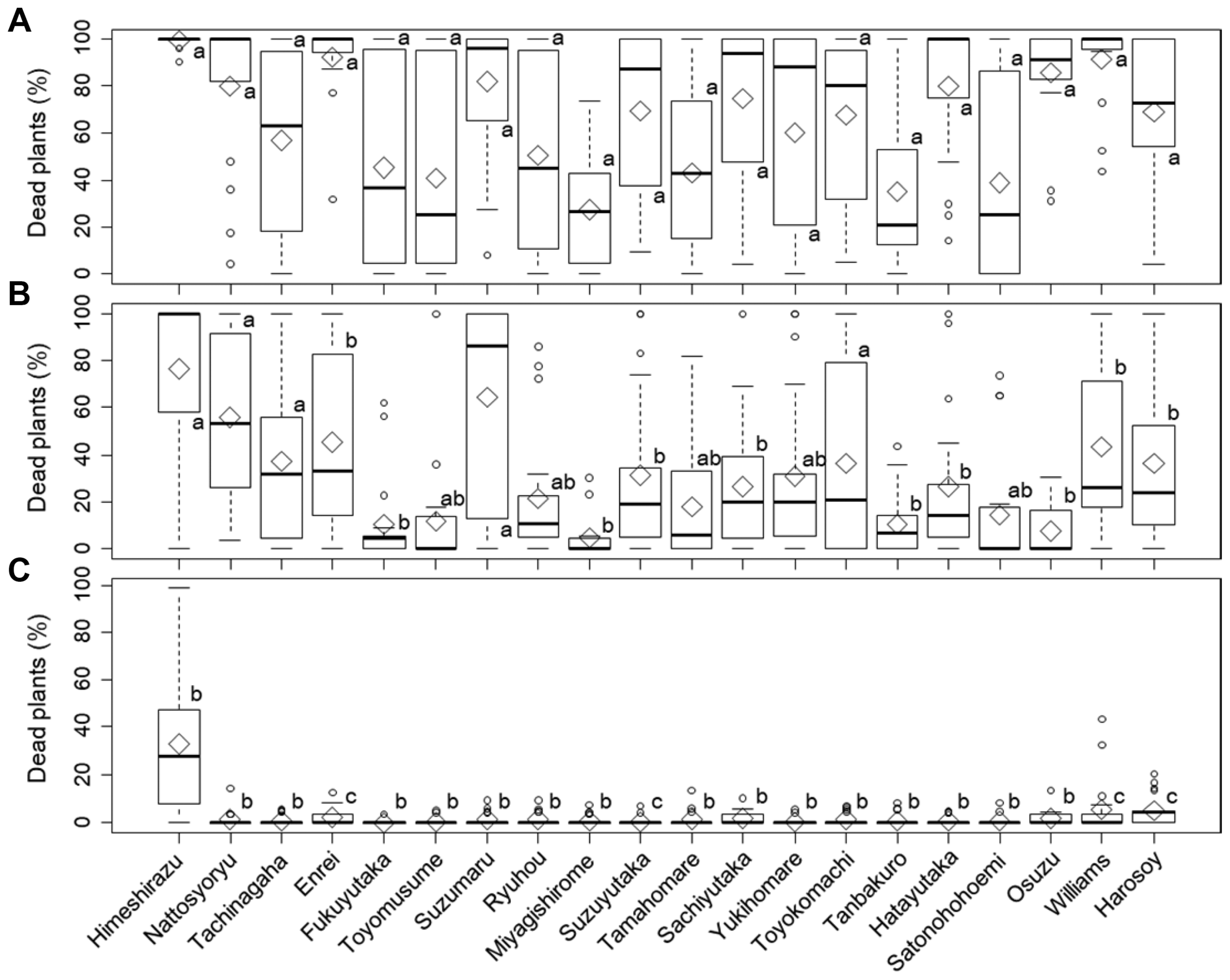
In a preliminary inoculation test involving 20 soybean cultivars, ‘Himeshirazu’ and ‘Nattosyoryu’ appeared to be susceptible to P. sojae isolate 13-B-RO-1. The average rates of dead plants for ‘Himeshirazu’ and ‘Nattosyoryu’ were greater than 90% when these cultivars were inoculated at 14 days after seeding or earlier. None of the control soybean plants treated with only V8 juice agar or water developed disease symptoms (Fig. 1). To investigate the temporal changes in the disease resistance levels of the two cultivars, plants grown for 16-40 days after sowing were inoculated with P. sojae isolate 13-B-RO-1 (Fig. 2). On average, more than 75% of ‘Himeshirazu’ and ‘Nattosyoryu’ plants died following the inoculations at 16-23 days and 16-18 days after sowing, respectively. Compared with the data for the plants inoculated at highly susceptible ages, the rates of dead plants for ‘Himeshirazu’ and ‘Nattosyoryu’ significantly decreased when plants were inoculated at 25-40 days and 26-40 days after sowing, respectively (Tukey’s HSD test, p < 0.05). The average rates of dead plants were less than 16% in ‘Himeshirazu’ plants grown for 29-40 days, and less than 5% in ‘Nattosyoryu’ plants grown for 29-40 days. To investigate whether the rates of dead plants decreased in other Japanese cultivars, the following 18 cultivars were challenged with 17 P. sojae isolates from Japanese soybean fields (Table 1) at 14, 21, and 28 days after seeding (Fig. 3): ‘Himeshirazu’, ‘Nattosyoryu’, ‘Tachinagaha’, ‘Enrei’, ‘Fukuyutaka’, ‘Toyomusume’, ‘Suzumaru’, ‘Ryuhou’, ‘Miyagishirome’, ‘Suzuyutaka’, ‘Tamahomare’, ‘Sachiyutaka’, ‘Yukihomare’, ‘Toyokomachi’, ‘Tanbakuro’, ‘Hatayutaka’, ‘Satonohohoemi’, and ‘Osuzu’ (Table 2). Two US cultivars, ‘Williams’ and ‘Harosoy’, were also inoculated with the same isolates to investigate their responses to Japanese P. sojae isolates (Fig. 3). Inoculations at 14 days after seeding resulted in an average rate of dead plants that exceeded 75% for the following six Japanese cultivars: ‘Himeshirazu’, ‘Nattosyoryu’, ‘Enrei’, ‘Suzumaru’, ‘Hatayutaka’, and ‘Osuzu’. The average rate of dead plants ranged from 27.7% to 74.8% for the 12 other Japanese cultivars. The extent of the resistance to the 17 P. sojae isolates varied significantly among the 20 cultivars (ANOVA, p < 0.001).
The average rate of dead plants decreased as the age of the plants when inoculated increased to 21 and 28 days after seeding. Moreover, there were significant differences in the extent of the resistance among cultivars for each age selected for inoculations (ANOVA, p < 0.001). During the inoculation of plants at 21 days after seeding, ‘Himeshirazu’ was the only cultivar with an average rate of dead plants that was greater than 75%. In contrast, ‘Nattosyoryu’, ‘Enrei’, and ‘Suzumaru’, which were also highly susceptible to disease when plants were inoculated at 14 days after seeding, had average rates of dead plants between 45.6% and 64.5%. ‘Hatayutaka’ and ‘Osuzu’ had even lower average rates of dead plants, at 26.4% and 7.8%, respectively. Of the plants inoculated at 14 days after seeding, only ‘Miyagishirome’ had an average rate of dead plants less than 30%. Meanwhile, during the inoculations at 21 days after seeding, the following 10 cultivars had an average rate of dead plants of 4.7%-26.4%: ‘Fukuyutaka’, ‘Toyomusume’, ‘Ryuhou’, ‘Miyagishirome’, ‘Tamahomare’, ‘Sachiyutaka’, ‘Yukihomare’, ‘Tanbakuro’, ‘Satonohohoemi’, and ‘Osuzu’. There were no significant differences in the rates of dead plants following the inoculations at 14 and 21 days after seeding for ‘Himeshirazu’, ‘Nattosyoryu’, ‘Tachinagaha’, ‘Toyomusume’, ‘Suzumaru’, ‘Ryuhou’, ‘Tamahomare’, ‘Yukihomare’, ‘Toyokomachi’, and ‘Satonohohoemi’. For the other cultivars, the rates of dead plants were significantly lower for the inoculations at 21 days after seeding (Tukey’s HSD test, p < 0.01). Of the plants grown for 28 days before being inoculated, only ‘Himeshirazu’ exhibited a significantly decreased degree of resistance against the P. sojae isolates (Tukey’s HSD test, p < 0.01). The average rate of dead plants for ‘Himeshirazu’ was 32.3%, which was considerably higher than the corresponding rate for the 17 other cultivars (0.2-2.5%). Moreover, for all cultivars, the average rates of dead plants were lower for the inoculations at 28 days after seeding than for the inoculations at 14 days after seeding (Tukey’s HSD test, p < 0.01). Thus, the inoculation test involving 18 Japanese cultivars and 17 P. sojae isolates revealed that the disease responses of the cultivars differed depending on the isolate used and the plant ages when inoculated.
The average rates of dead plants for the US cultivars, ‘Williams’ and ‘Harosoy’, were 91.2% and 69.1%, respectively, following the inoculation at 14 days after seeding. These averages for ‘Williams’ and ‘Harosoy’ decreased to 43.4% and 36.6%, respectively, following the inoculation at 21 days after seeding, and decreased further to 5.7% and 5.3%, respectively, following the inoculation at 28 days after seeding (Tukey’s HSD test, p < 0.01).
The results described herein indicated that the resistance levels against P. sojae isolates varied among the tested Japanese soybean cultivars. Additionally, none of the cultivars were completely resistant to all P. sojae isolates during the seedling and young plant stages. Moreover, for all cultivars, greater resistance (i.e., fewer dead plants) was observed with increasing plant age prior to inoculation. However, the plant ages at which the number of dead plants significantly decreased varied among the cultivars.
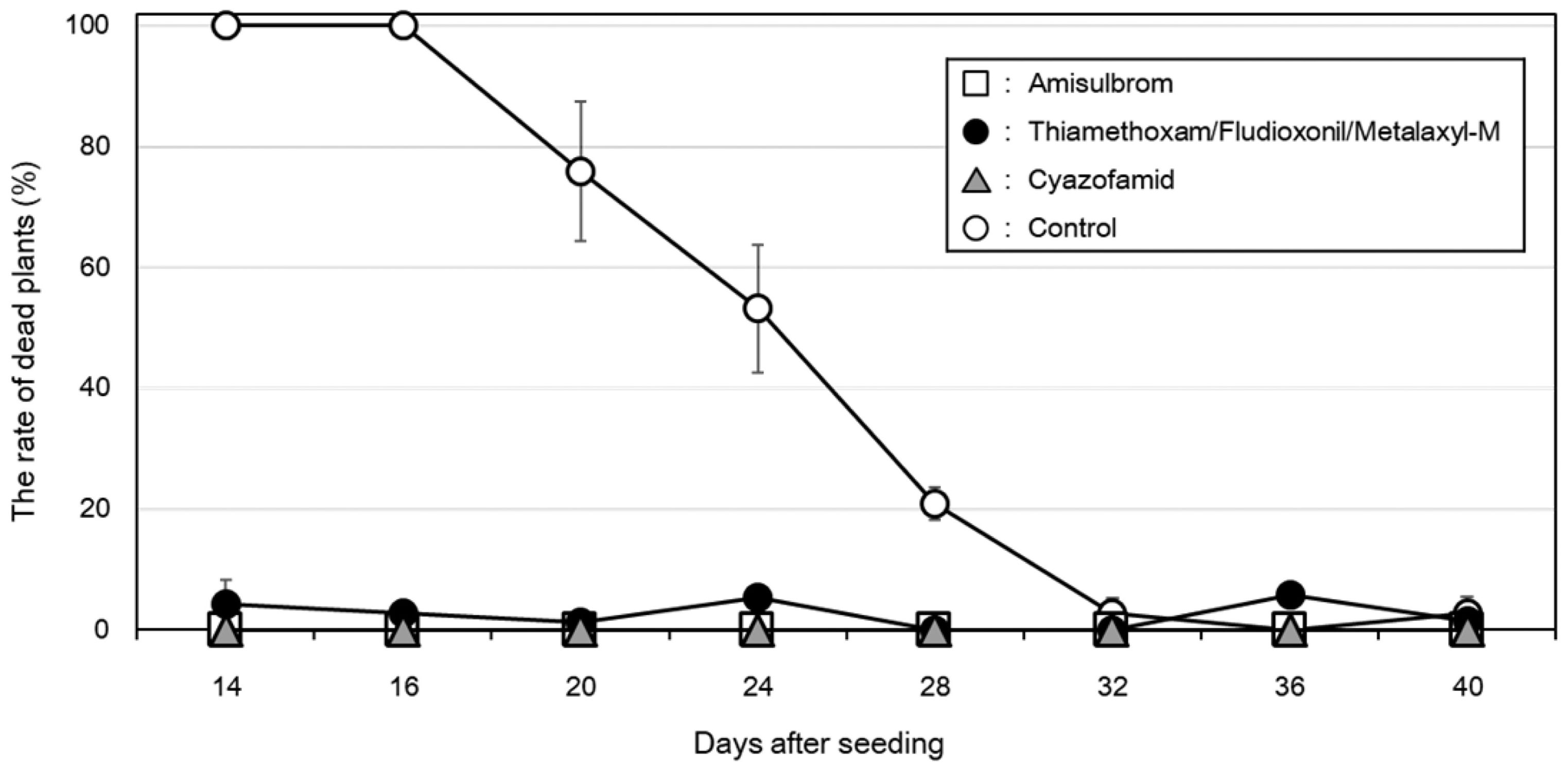
The effects of pesticide treatments of soybean seeds on the development of Phytophthora root and stem rot during the early soybean growth stages were investigated using three registered agrochemicals (Table 3, Fig. 4). The active ingredients against P. sojae in the three pesticides are amisulbrom, metalaxyl-M, and cyazofamid (Table 3). The rates of ‘Nattosyoryu’ plants killed by a P. sojae infection were significantly lower for the seed-treated samples than for the non-treated controls following the inoculations at 14, 16, 20, 24, and 28 days after seeding (Dunnett’s test, p < 0.001). For the inoculations at 32, 36, and 40 days after seeding, an average of ≤ 5.8% of ‘Nattosyoryu’ plants were killed regardless of whether seeds were treated with the pesticides. This result suggested that the three pesticides can control the pathogen during the growth period important for stand establishment. Additionally, the pesticide-induced decrease in the number of dead plants due to the pathogen was maintained for at least 28 days after treated seeds were sown.
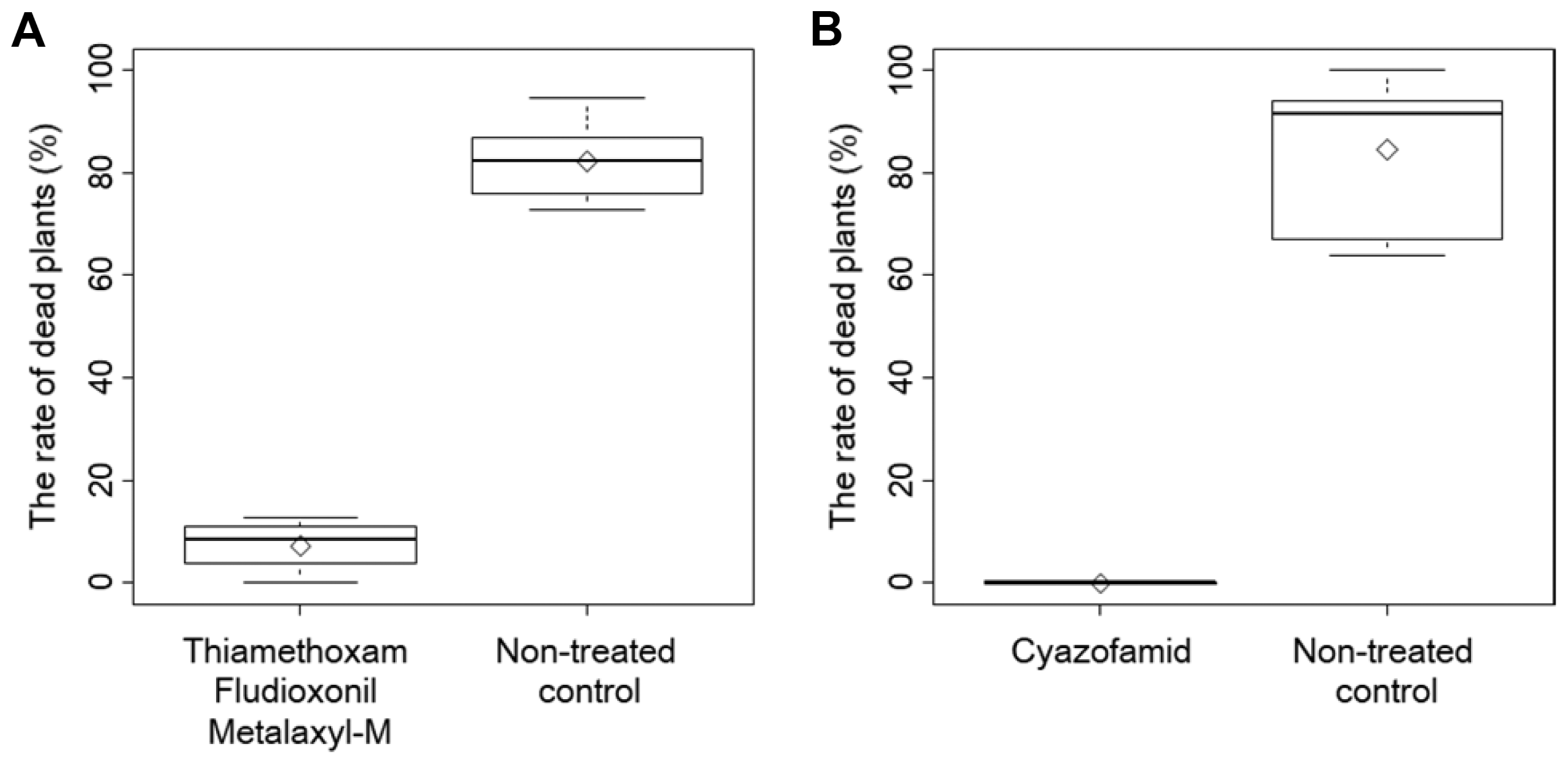
To investigate the efficacy of the pesticides against different P. sojae isolates, ‘Nattosyoryu’ plants were grown for 14 days after the seeds treated with the pesticides containing metalaxyl-M and cyazofamid were sown. The plants resulting from the seeds treated with metalaxyl-M were then challenged with the following seven P. sojae isolates: 13-B-RO-1, 13-B-RO-2, 13-7-B1-1, 13-8-11-2, Ps1001, Ps080806-3-5, and Ps060626-4-1. Meanwhile, the plants that emerged from seeds treated with cyazofamid were inoculated with the same P. sojae isolates, except for 13-B-RO-2. The number of dead plants significantly decreased in response to both pesticides (Fig. 5, Dunnett’s test, p < 0.05). The average rate of dead plants for the nontreated controls was 72.7%-100%, while the corresponding rate for the samples treated with metalaxyl-M was 0%-12.7%. Regarding the cyazofamid-treated samples, under the conditions in which an average of 63.7%-100% non-treated control plants were killed by the six P. sojae isolates, all of the pesticide-treated samples survived the inoculations.
Discussion
Phytophthora root and stem rot of soybean has threatened Japanese soybean production, especially when P. sojae infections occur during the early soybean growth stages. Soybean cultivation in Japan mainly involves the use of converted rice paddy fields. However, such a crop rotation system is favorable for P. sojae infections and disease development because of the soil type and poor drainage of the converted fields (Kato et al., 2013; Sugimoto et al., 2012). Vertical resistance via the expression of Rps genes provides complete protection from the corresponding pathogen races/populations (Dorrance, 2018), but Rps genes have not been exploited in Japan. Thus, the onset of Phytophthora root and stem rot during the early part of the soybean growing season leads to seed rot and seedling damping-off, with adverse consequences for stand establishment (Kato et al., 2013; Schmitthenner and Dorrance, 2015). In this study, we assessed the disease resistance of Japanese soybean cultivars, including the most widely grown cultivars in terms of planted area, by inoculating the aboveground part of the lower hypocotyl. Although none of the Japanese soybean cultivars were totally resistant to all tested P. sojae isolates during the first month after seeding, all cultivars exhibited increased resistance when plants were grown for more than 14 days after seeding before being inoculated. Additionally, a seed treatment with one of three pesticides protected soybean plants from P. sojae during the early vegetative growth phase, which is an important period for stand establishment. The protective effects of the pesticides were observed for at least 28 days after the treated seeds were sown. In the absence of strong resistance, like that mediated by Rps gene expression, in soybean seeds or seedlings immediately after germination, certain practices (e.g., seed treatments with agrochemicals) are needed to protect soybean plants from P. sojae infections during the early growth stages.
In this study, we evaluated the resistance of Japanese soybean cultivars to Phytophthora root and stem rot using a P. sojae-V8 culture flooding method modified from the procedure described by Mukobata and Sekihara (2005). Diverse inoculation methods have been developed, including the hypocotyl inoculation, greenhouse inoculum-layer test, and slant-board test as well as the application of a zoospore suspension and a mixture of agar culture homogenate and soil (Dorrance et al., 2004b, 2008; Dorrance and Mc-Clure, 2001; Guérin et al., 2014; Jiang et al., 2017; Stewart and Robertson, 2012; Sugimoto et al., 2006; Tooley and Grau, 1982). Some of these methods are used to evaluate complete resistance (e.g., Rps-mediated resistance), while others have been designed to assess incomplete resistance. In our method, the P. sojae agar culture is directly added to the water flooding the vermiculite in pots. Zoospores from the sporangia in the water initiate the infection, likely reproducing the soybean-pathogen interactions in the aboveground part of the lower hypocotyl when soybean plants are waterlogged during the early part of the growing season. This inoculation method enables P. sojae zoospores to mainly infect the aboveground part of the lower hypocotyl because negative geotaxis causes the zoospores to accumulate near the water surface (Cameron and Carlile, 1977). The damage to the lower hypocotyl caused by P. sojae severely inhibits soybean growth because of poor transport of water and/or nutrients from the roots to the upper plant parts, which leads to the damping-off of soybean seedlings and young plants. Our simple inoculation method does not require special techniques or expertise and results in the consistent induction of disease symptoms in soybean plants during the early vegetative growth phase. Moreover, this inoculation method revealed that the tested Japanese cultivars are differentially resistant to P. sojae. Although there is relatively little available information regarding the presence/absence of Rps genes in Japanese soybean cultivars (Athow et al., 1980; Rennie et al., 1992; Sugimoto et al., 2011, 2012), some of the tested cultivars were highly resistant to certain P. sojae isolates, suggesting that such strong resistance responses may be attributed to a race-specific resistance via the expression of Rps genes. A published report recommended the use of the hypocotyl inoculation test to assess Rps-mediated resistance to ensure P. sojae isolates compatible with the tested Japanese soybean cultivars prior to an evaluation of incomplete resistance (Dorrance et al., 2008). Because Rps-mediated resistance was not the focus of our study, compatible/incompatible interactions between the 18 Japanese cultivars and the 17 P. sojae isolates were not assessed before applying the P. sojae-V8 culture flooding method to evaluate the resistance of Japanese cultivars to P. sojae at different plant ages.
Partial resistance cannot completely prevent pathogen multiplication, while tolerance refers to the ability of the plant host to endure the presence of the pathogen (Parlevliet, 1979). In soybean, both resistance types are equivalently applied along with quantitative resistance, field resistance, and rate-reducing resistance (Dorrance, 2018; Schmitthenner, 1985). Tolerance to Phytophthora root and stem rot is defined as a relative characteristic based on the degree of plant loss or the decrease in growth or yield in the presence of the pathogen races for which a soybean cultivar lacks specific resistance genes (Anderson and Buzzell, 1982). Although it is unclear whether Japanese soybean cultivars carry certain Rps genes, we observed that the resistance of the tested soybean cultivars increased as the age of the plants when inoculated increased (i.e., decrease in the extent of plant death caused by the P. sojae infection). The inoculation of plants at 14 days after seeding revealed that six Japanese cultivars (‘Himeshirazu’, ‘Nattosyoryu’, ‘Enrei’, ‘Suzumaru’, ‘Hatayutaka’, and ‘Osuzu’) had an average rate of dead plants that exceeded 75%. With the exception of ‘Miyagishirome’, the other cultivars were also highly susceptible to certain P. sojae isolates. Thus, the resistance of the Japanese cultivars to P. sojae infections is not fully expressed in 14-day-old plants, in which the first trifoliate leaves are usually expanding or have expanded. Depending on the cultivars, the average rates of dead plants following a P. sojae infection significantly decreased when plants were inoculated at 21 or 28 days after seeding. Considering the partial resistance of soybean to Phytophthora root and stem rot is effective during or after the growth stage when the first trifoliate leaf forms (Dorrance and McClure, 2001; Schmitthenner and Dorrance, 2015), the plant age-dependent resistance observed in this study may be an example of partial resistance to the Japanese P. sojae isolates. Several plant species exhibit a disease resistance at various developmental stages, with varying levels depending on plant age or tissue maturity (Develey-Rivière and Galiana, 2007). The relationship between plant age/maturity and disease resistance has been studied in a soybean-P. sojae pathosystem (Dorrance and McClure, 2001; Jimenez and Lockwood, 1980; Lazarovits et al., 1981; Paxton and Chamberlain, 1969). Consistent with the results described herein, soybean plants reportedly become more resistant to P. sojae as they mature. For example, a previous study revealed that nine susceptible cultivars grown for 7 days before being inoculated were killed by P. sojae at a rate of 90% or higher, while the same cultivars grown for 21 days before being inoculated died at a rate of 30% or lower (Paxton and Chamberlain, 1969). In the hypocotyl, newer tissues are more susceptible to P. sojae than older tissues (Lazarovits et al., 1981). Additionally, an inoculation of ‘Altona’ plants grown for 6 days with zoospores of an incompatible race 4 isolate reportedly induces a resistance reaction in all hypocotyl parts, implying that the Rps-mediated incompatible interaction has been established in 6-day-old hypocotyls. The observed diseased or dead plants resulting from the inoculation of Japanese soybean cultivars at 14 days after seeding, suggests there is a lack of strong Rps-mediated resistance in most cultivar-isolate interactions at this plant growth stage. Soybean cultivar ‘Conrad’, which does not carry Rps genes and expresses a high level of partial resistance (Fehr et al., 1989), is highly susceptible to P. sojae infections at 0-5 days after seeding, with a seedling emergence rate significantly lower than that of plants inoculated later (Dorrance and McClure, 2001). Jimenez and Lockwood (1980) observed that even ‘Hark’, which is a susceptible cultivar with little field tolerance, exhibits increased resistance to P. sojae when plants are inoculated at 16 days after seeding or later. In the current study, the rates of dead plants due to a P. sojae infection for two susceptible cultivars, ‘Himeshirazu’ and ‘Nattosyoryu’, were significantly lower for plants inoculated at 25 and 26 days after seeding or later than for plants inoculated at 16-21 days and 16-18 days after seeding, respectively. The resistance of these two cultivars may not be expressed as quickly as that of ‘Conrad’. Although different inoculation and evaluation protocols were used, disease resistance increased as the age of plants at inoculation increased for all of the tested Japanese soybean cultivars, which is consistent with the results of previous studies (Dorrance and McClure, 2001; Jimenez and Lockwood, 1980; Lazarovits et al., 1981; Paxton and Chamberlain, 1969). The plant ages associated with increases in resistance varied among the cultivars, but the timing of the expression of resistance in the Japanese cultivars is likely consistent with that of partial resistance (Schmitthenner and Dorrance, 2015). Varying levels of partial resistance to P. sojae were shown in ‘Conrad’ and ‘Sloan’ hypocotyls, while a lack of partial resistance was observed in the roots of these two cultivars (Yamamoto et al., unpublished). However, partial resistance to P. sojae is reportedly expressed in the roots of cultivars with high levels of resistance (i.e., limited root rot and the absence of stem rot) (Dorrance et al., 2007; Mideros et al., 2007; Schmitthenner, 1985; Tooley and Grau, 1984). The partial resistance expression site in the current study might be inconsistent with that of several other studies that assessed root damage (Abeysekara et al., 2016; Burnham et al., 2003; Jia and Kurle, 2008; Lee et al., 2013a, 2013b; McBlain et al., 1991; Schneider et al., 2016; Stewart and Robertson, 2012; Tucker et al., 2010; Wu et al., 2011). This discrepancy was likely because our evaluation was based on the damping-off elicited by the severe infection around the lower hypocotyl and not root damage.
This study also determined the effective period for the treatment of seeds with three pesticides to provide protection against P. sojae. The active ingredients against Phytophthora root and stem rot in these pesticides are amisulbrom, metalaxyl-M, and cyazofamid. Metalaxyl-M, which is also known as mefenoxam, is the active isomer of metalaxyl, and is grouped with the phenylamide fungicides (Hirooka and Ishii, 2013), which are among the most commonly applied agrochemicals worldwide to control diverse oomycete-induced diseases (Dorrance, 2018; Dorrance et al., 2004a, 2007; Matheron and Porchas, 2014; Schmitthenner and Dorrance, 2015; Taylor et al., 2002). Meanwhile, amisulbrom and cyazofamid are quinone inside inhibitors that have similar protective effects against foliar and tuber blight caused by Phytophthora infestans (Hirooka and Ishii, 2013; Honda et al., 2007). Cyazofamid strongly inhibits the mycelial growth of oomycete species, with 0.2 μg/ml representing an effective concentration for inhibiting P. sojae growth by 50% (Mitani et al., 2001). At least 0.5 μg/ml is needed for an inhibition exceeding 90%, similar to metalaxyl. Additionally, mefenoxam is more effective than cyazofamid against Phytophthora capsici, with lesion development inhibited for at least 3 weeks when applied to roots (Matheron and Porchas, 2014). Although the three active ingredients are highly specific for oomycetes, previous studies focused primarily on the protective effects of metalaxyl and metalaxyl-M against Phytophthora root and stem rot of soybean (Anderson and Buzzell, 1982; Dorrance and McClure, 2001; Dorrance et al., 2009; Guy et al., 1989; Vaartaja et al., 1979). There have been relatively few studies examining the effects of amisulbrom and cyazofamid on soybean-P. sojae interactions (Mimuro, 2011). The current study revealed that the rates of dead plants due to P. sojae infections decreased significantly after seeds were treated with the pesticides. The protection provided by the pesticides continued for 28 days after the treatment. Because few plants were killed in the non-treated control at 29 days or later, the effective periods for these pesticides were not determined in this study. Mimuro (2011) reported that these three pesticides can protect soybean plants from Phytophthora root and stem rot for approximately 1 month after treated seeds are sown, which is consistent with our findings as well as the effective period indicated by the manufacturer of the pesticide containing metalaxyl-M.
According to industry estimates, the treatment of soybean seeds with pesticides has dramatically increased from < 8% in 1996 to > 30% in 2008, which has improved disease management during the early part of the soybean growing season (Esker and Conley, 2012; Munkvold, 2009). Soybean cultivation in Japan has recently become increasingly dependent on seed treatments, mainly with metalaxyl-M-containing pesticides. This is because soybean plants grown in fields converted from rice paddies are at high risk of being infected by P. sojae. Although seed treatments with pesticides are important for protecting young soybean plants from P. sojae and enhancing stand establishment, the pesticides are costly and repeated applications of the same pesticide may decrease the sensitivity of P. sojae to the active ingredient (i.e., pesticide resistance) (Han et al., 2011; Hunger et al., 1982). Planting cultivars exhibiting complete and/or incomplete disease resistance may be a more economically viable option for combating Phytophthora root and stem rot of soybean. Consequently, a comprehensive characterization of the disease resistance in Japanese cultivars via an analysis of Rps genes may be important for elucidating the Japanese soybean-P. sojae pathosystem and may be useful for the development of new lines/cultivars that are resistant to Phytophthora root and stem rot. The fact that none of the 18 tested Japanese cultivars were completely resistant to all of the P. sojae isolates following inoculations at 14 days after seeding suggests these cultivars likely do not express effective resistance during the early growth stages. Thus, additional practices, such as seed treatments with agrochemicals, will need to be implemented, especially if these soybean cultivars lack Rps-mediated resistance and are grown in disease-conducive environments, including fields previously infected by P. sojae and fields that have been waterlogged for prolonged periods.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






