

Biocontrol using microbes derived from natural sources such as soil, water, plants, and other organisms is one of the significant strategies for ecologically sound plant disease management. Disease control by antagonistic microbes can be achieved through antibiotic production, colonization, nutrient competition, induced systemic resistance (ISR), plant growth promotion, and/or parasitism against target plant pathogens (Ahmadzadeh and Tehrani, 2009; Choudhary and Johri, 2008; Kim et al., 2012; Kloepper et al., 2004; Perazzolli et al., 2008; Sang et al., 2011; Zhang et al., 2010). Despite many attempts to develop successful biocontrol microbes (agents), these agents often do not show consistent disease suppression relative to comparable commercial fungicides. This inconsistency is often based on inappropriate screening procedures to select effective biocontrol agents, due to a lack of understanding of competition or mutualism in the niche around host plants and various environment changes. For the successful practical applications of biocontrol agents, screening procedures would need to consider interactions with potential biocontrol agents, pathogens, and host plants, thereby minimizing the labor and time needed to obtain effective biocontrol agents (Chang et al., 2001; Kim et al., 2008a; Landa et al., 2004; Maleki et al., 2011).
Pepper (Capsicum annuum L.) is one of the most important cash crops in the world, including in Korea; however, its production and quality have been mainly limited by plant diseases such as Phytophthora blight and anthracnose. Phytophthora blight caused by an oomycete pathogen, Phytophthora capsici, is a destructive disease in pepper plants. This pathogen has a wide host range with more than 50 plant species including Cucurbitaceae, Leguminosae, and Solanaceae (Hausbeck and Lamour, 2004; Hwang and Kim, 1995; Tian and Babadoost, 2004). This soilborne pathogen can rapidly spread to the surface water via zoospores, produce persistent propagules including sporangia, mycelia, and oospores in the debris of infected plants, and infect all parts of pepper plants such as root, crown, foliage, and fruit through water splash (French-Monar et al., 2006; Hausbeck and Lamour, 2004; Lamour and Hausbeck, 2003). For these reasons, the management of this disease has been considered difficult and mainly depended on the use of agricultural chemicals, which sometimes causes fungicide resistance of the pathogen and toxicity to the crops (Cohen and Coffey, 1986; Lamour and Hausbeck, 2000, 2003; Parra and Ristaino, 2001). Thus, biocontrol has been considered as an alternative strategy for disease management in this vegetable crop (Kim et al., 2008a, 2009; Sang and Kim 2011; Sang et al., 2007, 2008, 2010). In this regard, many bacterial strains belonging to the genus Bacillus, Pseudomonas, and Paenibacillus have been used as biocontrol microbes against various soilborne pathogens with diverse disease suppressive mechanisms as described above. Recently, an antagonistic rhizobacterial strain, Flavobacterium johnsoniae GSE09, was shown to produce a volatile compound (2,4-di-tert-butylphenol), which exhibited antioomycete activity against P. capsici in pepper plants (Sang and Kim, 2012a; Sang et al., 2011).
Along with Phytophthora blight, anthracnose caused by several Colletotrichum spp. (mainly Colletotrichum acutatum) is also considered as one of the most destructive diseases that limit pepper production and quality (Park and Kim, 1992; Peres et al., 2005; Sang et al., 2011). Many biocontrol microbes have been reported to be capable of reducing anthracnose occurrence on various plants including chilli pepper, cucumber, bean, and banana by either seed or foliar-spray treatment (Bardas et al., 2009; Chanchaichaovivat et al., 2008; Fu et al., 2010; Neher et al., 2009). In addition to the direct inhibitory effect of the biocontrol microbes by the spray treatment, they often show a suppressive efficacy through ISR or plant growth promotion when applied into the roots or rhizosphere. Thus, the selection of effective biocontrol microbes against these destructive diseases of pepper plants is critically important with considering their mode of action in the host-pathogen interaction.
In our previous study, we developed a sequential screening procedure to select effective biocontrol bacterial strains against P. capsici in pepper plants (Kim et al., 2008a). Therefore, in this study, we conducted (i) to select effective antagonistic strains against Phytophthora blight of pepper using the developed screening procedure and (ii) to evaluate the effect of the antagonistic strains selected from the screening procedure on Phytophthora blight and anthracnose occurrence and fruit yield in the plants under field and plastic house conditions from 2007 to 2009. In addition, we identified the selected bacterial strains using fatty acid-methyl ester (FAME) and 16S rRNA gene sequence analyses.
Materials and Methods
Isolation and preparation of bacterial strains
Bacterial strains were isolated from the rhizosphere soil, and from the interior and surface of roots of cucumber, pepper, and tomato plants grown in fields in two locations [Youngjongdo (37┬░48N. lat., 126┬░49ŌĆ▓W. long.) and Andong (36┬░56ŌĆ▓N. lat., 128┬░76ŌĆ▓W. long.)], Korea in 2004 and 2006. Soil-adhering plant roots were collected in polyethylene bags, stored in an ice chest, and used within 2-3 days after collection. To obtain bacterial strains from rhizosphere soil, the soil (10 g) was placed in 100 ml sterile water. For the isolation of strains from the root surface, roots (10 g) were used after the complete removal of soil. To isolate strains from the interior of root, roots (10 g) were surface-sterilized with 1% sodium hypochlorite for 90 sec, rinsed several times in sterile distilled water, and completely macerated with a sterile homogenizer. The macerated roots were placed in 100 ml sterile water and shaken at 160 rpm at 28┬░C for 30 min. The suspension (200 ╬╝l) was spread on tryptic soy agar (TSA) containing 50 ╬╝g/ml of cycloheximide and cultured at 28┬░C for 48 h. Distinct colonies appeared on TSA were selected, streaked on nutrient agar (NA), and incubated at 28┬░C for 48 h. From these cultures, single colonies were obtained and cultured in nutrient broth (NB) in a rotary shaking incubator (160 rpm) at 28┬░C for 24 h, and then stored in NB containing with 20% glycerol at ŌłÆ70┬░C until used.
For pepper radicle and seedling assays, bacterial strains were streaked on NA and incubated at 28┬░C for 48 h. The bacterial cells were harvested and suspended in 10 mM MgSO4 solution, and then adjusted to 108 cells/ml (OD600 = 0.5) with a spectrophotometer (Du┬« 650, Beckman Coulter, Fullerton, CA, USA). Likewise, for plant, field, and commercial plastic house tests, single colonies were inoculated in 5 ml NB and incubated in a shaking incubator (160 rpm) at 28┬░C for 24 h. The pre-cultured bacterial cells were further incubated in 500 ml NB in a shaking incubator (160 rpm) at 28┬░C for 48 h. The cells were harvested in 10 mM MgSO4 solution by centrifuging at 5,000 ├Ś g at 20┬░C for 15 min, and the harvested pellets (bacterial cells) were washed twice with the same solution by centrifugation. Bacterial suspensions were adjusted to 108 cells/ml in the solution as described above.
Radicle and seedling assays against P. capsici
For the radicle assay, uniformly germinated pepper (cv., Nockwang) seeds (2-3 mm in radicle) were soaked in the bacterial suspensions for 3 h and blotted on sterile filter papers. Ten seeds per bacterial strain were placed at 28┬░C in the margins of 5-day-old cultures of a virulent isolate S197 of P. capsici on water agar (WA) containing 0.02% glucose. These treated plates were incubated at 28┬░C under 16 h fluorescent light/day. Seeds treated with 10 mM MgSO4 solution were used as a control. Incidence (%) of infected radicles with brownish discoloration was determined 23 days after placing the seeds on WA, when almost all the control seeds were infected. The selected strains from the initial radicle assays were tested twice again with three replicates of 10 seeds each.
For the seedling assay, germinated pepper (ŌĆśNockwangŌĆÖ) seeds were planted in 128-cell (3 ├Ś 3 ├Ś 5 cm) plug trays filled with potting mix (Sang et al., 2008) that included the bacterial suspensions. The bacterial strains were incorporated at 108 cells/g dry weight of potting mix. The 10 mM MgSO4 solution was used as an untreated control. The trays were placed at room temperature in a growth room under 16 h fluorescent light/day. After 2 weeks, pepper seedlings were inoculated with P. capsici zoospore suspension (2 ├Ś 103 zoospores/g dry wt. potting mix) at 1 cm below the soil-line near the seedlings. The pathogen inoculum was prepared using the procedure described by Kim et al. (1997). Disease incidence (%) was evaluated 4 days after inoculation. The selected strains from the radicle assays were tested twice with three replicates of eight seedlings each.
In planta trials against P. capsici.
Eleven bacterial strains, which were selected from the radicle and seedling assays, were tested in 5-week-old pepper (ŌĆśNockwangŌĆÖ) plants in a growth room. Pepper seedlings, bacterial suspensions, and pathogen inocula prepared as described above were used in the in planta trial. Three-week-old seedlings grown in the potting mix were transplanted into steam-sterilized soil in 10 cm-diameter pots with saucers. One week later, the pots were drenched with 25 ml of each bacterial suspension. Additionally, a week later, these plants were inoculated with P. capsici zoospores (25 zoospores/g dry wt. soil) as follows: zoospores were injected into four holes (1 cm diameter ├Ś 1 cm deep) around each plant. These inoculated plants were continuously watered for 4-5 days through the saucers to prevent drying of the soil, and then watered as needed. Inoculated and uninoculated plants drenched with 10 mM MgSO4 solution were used as positive and negative controls, respectively. Disease severity, on a scale of 0 (symptomless) to 5 (plants dead) as described by Kim et al. (2012), was evaluated 17 and 18 days after inoculation in repeated experiments. The experiment was performed twice in a completely random design with 15 plants each.
Field tests for Phytophthora blight and pepper fruit yield
Field tests with the antagonistic bacterial strains (YJR27, YJR92, YJR102, and YJR107) selected from in planta trials were conducted at the Deokso Experiment Farm of Korea University, Namyangju, Korea from 2007 to 2009. Tests were established in beds with 7.5 m-long plots for the 2007 test and 12.5 m-long plots for the 2008 and 2009 tests, and arranged in a randomized complete block with three replicates per treatment for the 2007 test and four replicates per treatment for the 2008 and 2009 tests. Raised beds (20 cm high ├Ś 50 cm wide), spaced 85 cm apart (center to center), were constructed and covered with black plastic mulch. Pepper plants (ŌĆśNockwangŌĆÖ) were grown for 10 weeks in 2007, 13 weeks in 2008 and 11 weeks in 2009 in pots containing potting mix (Sang et al., 2008). The plant roots were dipped in one of the following treatments: each of the four bacterial suspensions, 10-mM MgSO4 solution (untreated control), or metalaxyl (a.i. 7.5%, Ridomil MG┬«, Dongbu Hannong Chemicals, Seoul, Korea) (fungicide control). These treated plants were transplanted into the beds in rows (30 cm between plants) on 1 June 2007, 16 June 2008 and 14 May 2009. Next, 10 ml of P. capsici (104 zoospores/ml) was inoculated into the soil around each plant on 13 August 2007, 11 August 2008 and 22 July 2009. Disease incidence (%) and severity, as described above, were regularly evaluated after the first appearance of symptoms on the inoculated plants.
To assess root infection by P. capsici, three plant roots per plot were collected on 17 September 2007, 10 September 2008, and 2 September 2009. The soil attached to the collected roots was removed manually and the roots were cut into 1 cm-long fragments. The 100 root fragments (three sub-replicates per root) were placed on pimaricin-ampicillin-rifampicin-pentachloronitrobenzene-hymexazol (PARPH) medium (Solel and Pinkas, 1984) selective for Phytophthora spp. and incubated at 28┬░C. The numbers of root fragments infected by P. capsici were evaluated 3-4 days after placing on the medium. The experiment was conducted with three replicates of three plant roots each in 2007 and four replicates of three plant roots each in 2008 and 2009. The same root samples collected for root infection assessment were used to evaluate total populations of bacteria and fungi. The rhizosphere soil (10 g) of each plant (three sub-replicates per plant) were placed in 100 ml sterile water and shaken at 160 rpm at 28┬░C for 30 min. The suspension was spread and cultured at 28┬░C for 48 h on g cycloheximide/ml for total bacteria TSA containing 50 ╬╝ and Ohio Agricultural Experiment Station (OAES) medium (Williams and Schmitthenner, 1960) for total fungi. These experiments were conducted with the same replicates used in the root infection assessment. Bacterial and fungal colonies were counted 2 and 3 days after incubation, respectively, and assessed based on the dry weight of soil.
Pepper fruit production was evaluated to assess the plant growth-promoting effect by the selected bacteria in the 2007 and 2008 tests. Numbers and fresh weight (kg) of marketable pepper fruits (> 8 cm long), both red (ripe) and green (unripe), were counted four times after harvesting, on August 6 and 13, and September 8 and 22 in 2007, and twice on August 12 and 21 in 2008. These experiments were conducted with three replicates of 25 plants each in the 2007 test and four replicates of 40 plants each in the 2008 test.
Plastic house tests for anthracnose and pepper fruit yield
Anthracnose and fruit yield affected by the same selected bacterial strains were evaluated in a commercial, plastic (polyethylene; PE)-covered house (18 m wide ├Ś 45 m long) at the same University Farm in 2008 and 2009. Tests were arranged in beds with 3.6 and 4.5 m-long plots (two beds per treatment) and designed in a randomized complete block with five and four replicates per treatment in the 2008 and 2009 tests, respectively. Raised beds (20 cm high ├Ś 80 cm wide), spaced 1 m apart (center to center), were covered with black plastic mulch. Pepper plants (cv., Buja) were grown for 10 weeks in 2008 and 11 weeks in 2009 in 10 cm-diameter pots containing potting mix (Sang et al., 2008). These plants in pots were drenched once with 100 ml of each bacterial strain suspension or 10 mM MgSO4 solution (untreated control) per pot on 19 April 2008 and 18 April 2009. After 2 (2009) or 3 (2008) days, plants were transplanted into the beds in rows (45 cm between plants). For pepper yield assessment, the numbers and fresh weights of marketable pepper fruits (> 8 cm long) were determined. Numbers of green (unripe) and red (ripe) fruits were counted twice, on August 2 and 25 in 2008 and on July 29 and August 19 in 2009, 2-5 days before harvesting the pepper fruits. The pepper fruit weights (red fruits only) were determined twice after harvesting, on August 4 and 27 in 2008 and on August 3 and 21 in 2009 for marketing purposes. To assess the effects of the bacterial strains against anthracnose by C. acutatum on pepper fruit, naturally infected green and red fruits with distinct symptoms were simultaneously evaluated at the time of fruit number evaluation. Disease incidence (%) on either the green or red fruit was determined as follows: number of infected green or red fruits divided by the total number of green or red fruits ├Ś 100. Numbers and fresh weights (kg) of pepper fruits, and anthracnose incidence were determined with five replicates of 15 plants each in the 2008 test and four replicates of 20 plants each in the 2009 test.
Identification of selected antagonistic bacterial strains
Total cellular FAME analysis of the selected antagonistic strains (YJR27, YJR92, YJR102, and YJR107) was conducted by gas chromatography using the Microbial Identification System (MIDI; Newark, DE, USA) according to the manufacturerŌĆÖs instructions. For 16S rRNA gene sequence analysis, the genomic DNA of the strains was isolated using the i-genomic BYF DNA Extraction Mini Kit (iNtRON Biotechnology, Seongnam, Korea), and the amplicon of the 16S rRNA gene was obtained by polymerase chain reaction (PCR) using the universal primers fD1 and rP2 (Kim et al., 2012). 16S rRNA gene sequence analysis was conducted using the BLAST network service at the National Center for Biotechnology Information (NCBI) of the U.S. National Library of Medicine (Bethesda, MD, USA). Phylogenetic trees were constructed using the neighbor-joining method and bootstrap analysis using Molecular Evolutionary Genetics Analysis (MEGA) software, version 5.05 (The Biodesign Institute, Tempe, AZ, USA) (Saitou and Nei, 1987).
Statistical analysis
Statistical analysis of data was conducted using the Statistical Analysis Systems (SAS Institute, Cary, NC, USA). Data from repeated experiments in each test were pooled after confirming the homogeneity of variances with LeveneŌĆÖs test (1960) and were further statistically analyzed. Percent data of disease incidence were statistically analyzed after arcsine square-root transformation; however, untransformed data are presented. The numbers of total bacteria and fungi in rhizosphere soils were analyzed after log transformation. For the analysis of ordinal data such as disease severity, nonparametric analysis was used based on the ranks of the data, but untransformed data are presented. Analysis of variance was determined using the general linear model procedures, and means were separated with the least significant difference (LSD) test at P < 0.05.
Results
Isolation of bacterial strains
A total of 239 bacterial strains were isolated from the surface or interior of the roots and from the rhizosphere soil of cucumber, pepper, and tomato plants grown in the fields in Andong and Youngjongdo, Korea in 2004 and 2006 (Table 1).
Selection of antagonistic bacterial strains against P. capsici using a sequential screening procedure
The 239 bacterial strains showed the various levels of protection of pepper radicles against P. capsici. In our previous study (Sang et al., 2008), we selected strains with at least 30% radicle protection to have a better possibility of obtaining effective antagonistic strains. In this regard, 11 bacterial strains (Table 2) among 239 strains tested were selected as having Ōēż70% radicle infection (Ōēź30% disease protection) by P. capsici. Among the 11 strains, four strains (YJR13, YJR27, YJR92, and YJR102) resulted in 50-60% radical infection (40-50% disease protection), and 7 strains (YJR03, YJR07, YJR59, YJR89, YJR96, YJR100, and YJR107) showed 60-70% radicle infection (30-40% disease protection) (Table 2). However, the other 228 strains showed little or no radicle protection against pathogen infection.
The 11 strains selected in this radicle assay, which were obtained from the root surfaces of pepper plants, were tested in the seedling assay. In the seedling assay, all these strains significantly (P < 0.05) reduced disease incidence by P. capsici, compared with MgSO4 treatment (untreated control) (Table 2). These 11 strains were then examined in the in planta trial, in which nine strains, with exception of strains YJR07 and YJR89, significantly (P < 0.05) suppressed disease severity compared with untreated controls. Consequently, four strains (YJR27, YJR92, YJR102, and YJR107) were selected as antagonistic strains for further field tests (Table 2).
Effect of rhizobacteria on Phytophthora blight occurrence and pepper fruit yield in the field plants with artificial inoculum
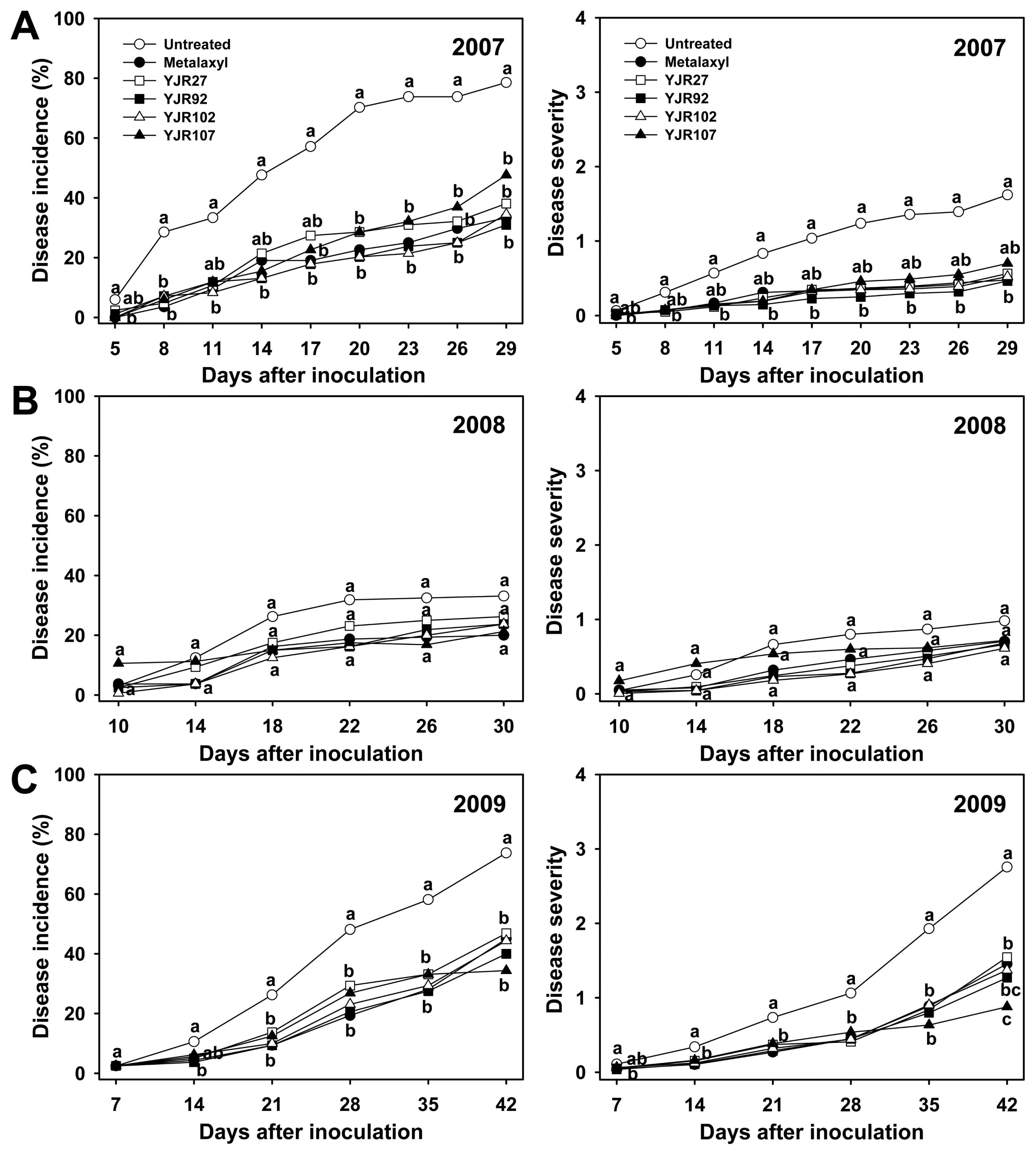
Phytophthora blight of pepper occurred severely in 2007 and 2009, and moderately in 2008, in the field plants that were artificially inoculated with P. capsici (Fig. 1). The final disease incidence and severity ranges were 31-77% and 0.5-1.6 in 2007, 20-33% and 0.6-1.0 in 2008, and 34-74% and 0.9-2.8 in 2009, respectively. In the 2007 test, all tested strains, with exception of strain YJR107 for disease severity, significantly (P < 0.05) reduced disease incidence and severity, compared with MgSO4 treatment (untreated control) across evaluation dates after inoculation (Fig. 1A). Strains YJR27, YJR92, and YJR102 protected pepper plants against pathogen infection at similar levels as observed in metalaxyl treatment (fungicide control). The MgSO4-treated plants produced the highest disease (disease incidence = 77% and disease severity = 1.6 at final evaluation) among all the treatments (Fig. 1A). However, in the 2008 test, moderate disease levels were observed, and none of the treatments were significantly different from untreated controls across disease evaluation dates (Fig. 1B). In the 2009 test, similar results were obtained as observed in the 2007 test (Fig. 1C). In the 2009 test, all tested strains significantly (P < 0.05) reduced disease incidence and severity compared with untreated controls across disease evaluation dates, and showed similar or better levels in disease suppression than in metalaxyl treatment (Fig. 1C).
As a parameter of disease infection by P. capsici, root infection rates were assessed in field tests. In the 2007 test, all the tested strains significantly (P < 0.05) reduced root infection in the range of 56-64%, compared with MgSO4-treated plants (untreated control) (Table 3). The results of root infection in the 2009 test were similar to those of the 2007 test; all tested strains reduced 43-62% of root infection, compared with untreated control. However, in the 2008 test, the root infection rates in all the bacterial and metalaxyl (fungicide control) treatments did not differ from those in the untreated controls. The root infection rates of MgSO4-treated controls in the 2008 test were lower than those in the 2007 and 2009 tests (Table 3). When total numbers of bacteria and fungi in the rhizosphere soils were compared, the populations of total bacteria per gram of dry soil ranged from 6.29-6.43 (log value) in 2007, 7.67-7.83 in 2008, and 7.31-7.99 in 2009 (Table 4). Likewise, populations of total fungi ranged from 3.11-3.26 (log value) in 2007, 4.10-4.23 in 2008, and 4.00-4.71 in 2009. Populations of total bacteria and fungi in bacteria-treated pepper rhizospheres were not significantly (P > 0.05) different from those in MgSO4- and metalaxyl-treated plants in test years (Table 4).
The numbers and fresh weights of pepper fruits were determined in the fields over 2 years (Tables 5 and 6). In the 2007 test, none of the tested strains yielded significantly (P > 0.05) different results compared with MgSO4 treatment (untreated control) in terms of fruit numbers at the first and second harvests (pre-inoculation) (Table 5). However, strains YJR27, YJR92, and YJR102 increased the fruit numbers at the third and fourth harvest (post-inoculation), compared with those of MgSO4-treated controls. Metalaxyl treatment increased the numbers at the post-inoculated harvests. Similar results were obtained with regard to fruit weights. Strains YJR27, YJR92, and YJR102 as well as metalaxyl increased fruit weights at the fourth harvest, compared with those of the untreated control. However, strain YJR107 reduced the numbers and weights of the pepper fruit at the first and second harvests (Table 5). In the 2008 test, the fruit numbers and weights of pepper plants treated with the bacterial strains and metalaxyl did not differ from MgSO4-treated controls (Table 6).
Effect of rhizobacteria on anthracnose occurrence and pepper fruit yield in the plastic house plants with natural inoculum
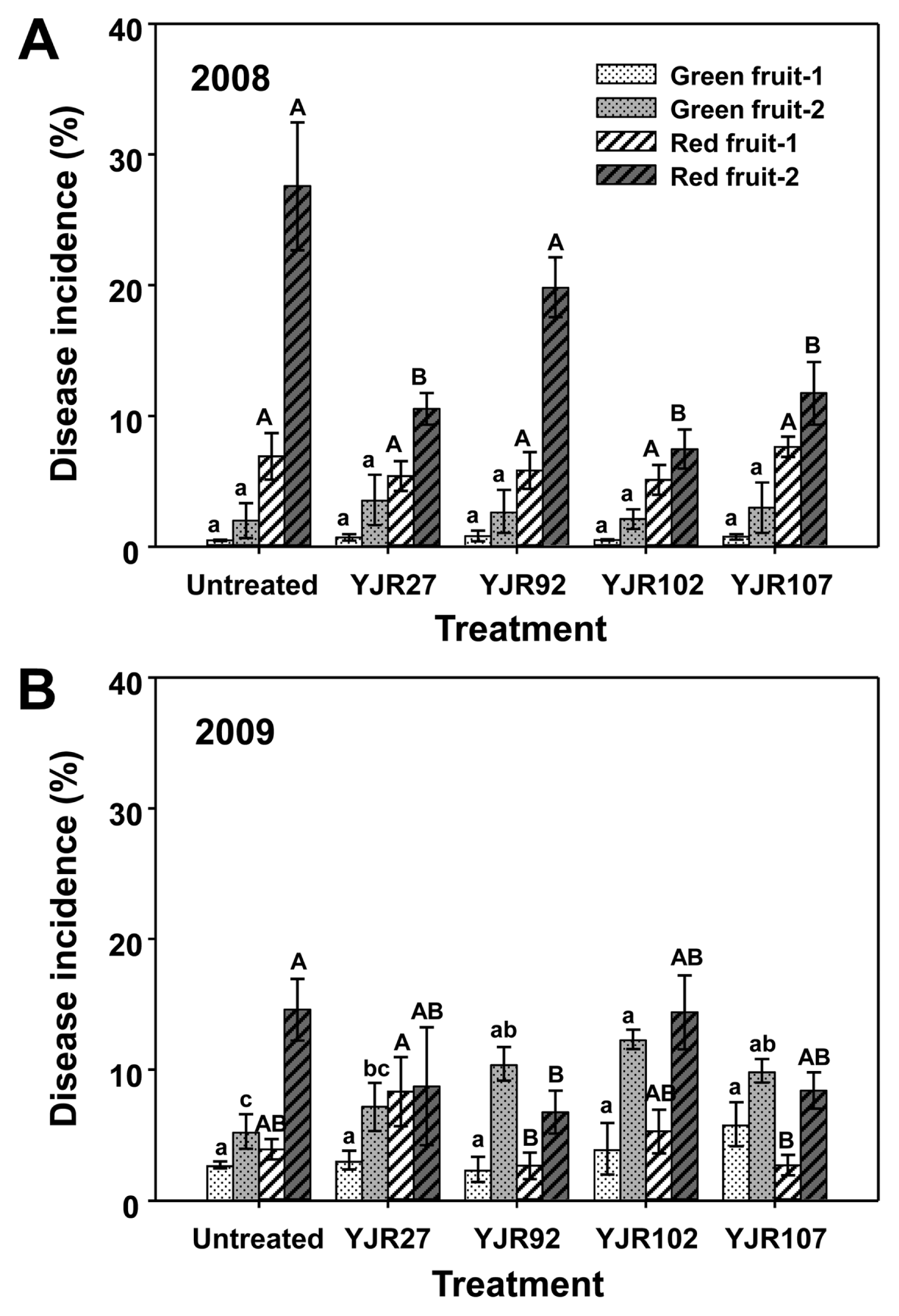
In the 2008 test, strains YJR27, YJR102, and YJR107, but not strain YJR92, significantly (P < 0.05) reduced anthracnose incidence (disease reduction rates = 57.4-73.0%) on red fruits at the second harvest, compared with MgSO4 treatment (untreated control) (Fig. 2A). However, all the tested strains failed to reduce disease incidence on green fruits at the first and second harvests, as well as on red fruits at the first harvest (Fig. 2A). In the 2009 test, strain YJR92 alone significantly (P < 0.05) reduced the disease incidence (disease reduction rate = 53.7%) on red fruit at the second harvest, compared with the untreated control (Fig. 2B). However, all the tested strains failed to reduce the disease incidence on green fruit at the first and second harvests, and on red fruit at the first harvest (Fig. 2B).
In the case of pepper fruit yield assessment, fruit numbers and fresh weights were affected by the bacterial strains in 2008 and 2009 (Table 7). In the 2008 test, strain YJR102 alone significantly (P < 0.05) increased the numbers of red fruit at the second harvest, compared with MgSO4 treatment. In terms of fresh weight of red fruit, strains YJR27 and YJR92 significantly (P < 0.05) increased the fruit weight at the first harvest. However, all strains failed to increase the numbers of green fruit at both the harvests and of red fruit at the first harvest, as well as the red fruit weight at second harvest. Similarly, as observed in the 2008 test, in the 2009 test, all the tested strains failed to affected the numbers of green and red fruits; however, three strains (YJR27, YJR92, and YJR102) significantly (P < 0.05) affected red fruit weights at the first harvest, but not at the second harvest (Table 7).
Identification of selected antagonistic bacterial strains
The four selected strains (YJR27, YJR92, YJR102, and YJR107) were identified using FAME and 16S rRNA gene sequence analyses. From the FAME profiles, major cellular fatty acids of strain YJR27 were C16:0 (22.9%), summed feature 3 (C16:1 Žē7c and/or C16:1 Žē6c) (20.3%), and C12:0 (12.1%) and those of strain YJR92 were C16:0 (30.6%), summed feature 3 (C16:1 Žē7c and/or C16:1 Žē6c) (20.2%), and summed feature 8 (C18:1 Žē7c) (18.5%) which identified the strain as belonging to the genus Pseudomonas. Strain YJR102 mainly had C18:1 Žē9c (34.7%), C16:0 (31.2%), and summed feature 3 (C16:1 Žē6c and/or C16:1 Žē7c) (17.2%), and was identified as belonging to the genus Tsukamurella. On the other hand, strain YJR107 had C18:1 Žē7c (57.7%) and C14:0 2OH (14.9%), and was identified as belonging to the genus Novosphingobium or related genus. Further, 16S rRNA gene sequence analysis identified strains YJR27 as Pseudomonas otitidis (similarity = 99%), YJR92 as Pseudomonas putida (99%), YJR102 as Tsukamurella tyrosinosolvens (100%), and YJR107 as Novosphingobium capsulatum (99%). The 16S rRNA gene sequences of strains YJR27 (accession number = JX154555), YJR92 (JX154556), YJR102 (JX154557), and YJR107 (JX154554) have been deposited in the GenBank.
Discussion
We previously described a sequential screening procedure requiring reduced time and labor, consisting of radicle and seedling assays and in planta trials, to select effective antagonistic bacterial strains against P. capsici in pepper plants (Kim et al., 2008a). In this study, we selected four strains from 239 tested strains as potential biocontrol agents against the pathogen using this screening procedure. Thereafter, we assessed the control efficacy of these selected strains against Phytophthora blight of pepper under field conditions with artificial inoculation from 2007 to 2009, and against anthracnose under plastic house conditions with natural inoculation in 2008 and 2009. Further, we evaluated the effect of these strains on pepper fruit yield under the same conditions. The results from this study confirmed that the developed screening procedure was reliable and rapid for selecting antagonistic bacterial strains against P. capsici in pepper plants. In addition, these selected strains exhibited biocontrol activities against anthracnose, and some of strains also exhibited plant growth-promotion effects on pepper fruit yield.
We selected four bacterial strains (P. otitidis YJR27, P. putida YJR92, T. tyrosinosolvens YJR102, and N. capsulatum YJR107) from the sequential screening procedure as potential biocontrol agents against the pepper disease. However, in selecting these strains in this study, we used a modified screening procedure (in terms of the baseline of the radicle assay and the inoculum concentration in the seedling assay) (Sang et al., 2008) from the initially developed procedure (Kim et al., 2008a) to obtain better antagonistic strains. For example, the initial screening baseline of the radicle assay was Ōēż 80% radicle infection (Ōēź20% radicle protection) but the baseline was increased to Ōēż 70% radicle infection (Ōēź 30% radicle protection) in this modified procedure. In addition, the inoculum concentration for the seedling assay was increased to 2 ├Ś 103 zoospores from 1 ├Ś 103 zoospores per gram dry weight of potting mix. From our previous study (Sang et al., 2008), we found that these severe selection pressures could significantly reduce the time and labor required to select appropriate antagonistic strains, and this was subsequently evaluated in in planta trials and field tests.
Further, we confirmed the merit of the modified procedure for selecting potential biocontrol strains. When comparing the initial baseline of 80% radicle infection (20% radicle protection) to the modified baseline of 70% (30% radicle protection) in the radicle assay, 88 of 231 strains (38.1%) were selected as potentially antagonistic strains using the 80% radicle infection in our initial study (Kim et al., 2008a). However, using the baseline of 70% radicle infection, 38 of 439 strains (8.4%) (Sang et al., 2008), and 11 of 239 strains (4.6%) in this study were selected as potentially antagonistic strains. Eventually, using the screening procedure, 11 of 231 strains (4.8%) (Kim et al., 2008a), 16 of 439 strains (3.6%) (Sang et al., 2008), and nine of 239 strains (3.8%) in this study were selected as antagonistic strains against P. capsici. In addition, these consistent results might be derived from the merit of the radicle assay as a pre-screening method. Since the radicle assay was designed to evaluate the interaction between the bacterial strain (antagonist), pepper radicle (host), and P. capsici (pathogen), strains from this assay have a good potential for survival and persist on the root surface or in the rhizosphere, which could play a significant role in disease suppression in the seedling assay and in planta trials. Recently, Barahona et al. (2011) indicated that the activity of bio-control agents could be improved by increasing their competitive colonization ability. Therefore, the sequential screening procedure based on the pre-screening radicle assay could offer not only rapid selection but also increased colonization ability of the selected strains against P. capsici in pepper plants.
In the 3-year field tests, the control efficacies of the selected strains (YJR27, YJR92, YJR102, and YJR107) against Phytophthora blight of pepper were examined. In the 2007 and 2009 tests, three strains YJR27, YJR92, and YJR102 exhibited significant biocontrol abilities against the disease; however, in the 2008 test, all the strains failed to reduce it. The disease reductions in the 2007 and 2009 tests were more obvious when observed in terms of root infection rates by P. capsici. The tested strains greatly reduced root infection by the pathogen relative to the untreated control, and showed similar efficacies as seen in plants treated with metalaxyl in the 2007 and 2009 tests, but not in the 2008 test. Our previous studies (Kim et al., 2009, 2012; Sang and Kim, 2012a) demonstrated that root protection through colonization by biocontrol strains against P. capsici could play an important role in disease suppression. However, in the 2008 test, the disease occurrence and root infection rates were not significantly different between the strain-treated plants and untreated or metalaxyl-treated control plants. The lack of significant difference in disease suppression might be due to low disease pressure in 2008, compared with severe disease pressure in the 2007 and 2009 tests; this could be affected by various environmental factors. Recently, Seo et al. (2011) reported that the occurrence of this pepper disease in the fields in Korea was lower in 2008 than in 2007. In addition, it was shown that the tested strains did not negatively affect the microbial population in the pepper rhizosphere when the influence of the strains on total bacterial and fungal populations was examined. It is desirable for the introduced biocontrol strains not to alter the microbial community or diversity in the environment. However, since only culturable bacteria and fungi on media were assessed in this study, further examination of the effect of these strains on the root and soil microbial community and diversity, including non-culturable microbes, is needed as studied by Sang and Kim (2012b).
In the 2-year plastic house tests, the tested same strains reduced anthracnose incidence by C. acutatum (Sang et al., 2011), possibly through ISR as observed in our previous works (Sang and Kim, 2011; Sang et al., 2011), at least one of test years but their biocontrol activities were variable. These variable results might be due to the limitations of the required populations of the treated strains for inducing effective ISR against anthracnose in pepper fruit. Recently, Nijhuis et al. (2010) suggested that a sufficient population of Lysobacter enzymogenes 3.1T8 on cucumber roots was required for the biocontrol of Pythium root rot. Consistency and improvement in the microbial biocontrol activity can be gained by the use of certain additional compounds (Shaukat and Siddiqui, 2003). Kim et al. (2008b) reported that the biocontrol activity of antagonistic Chryseobacterium sp. KJ1R5 against P. capsici could be enhanced by the addition of certain carbon sources. In another study, Postma et al. (2009) found that higher numbers of L. enzymogenes 3.1T8 were obtained by adding chitosan, resulting in effective control of Pythium aphanidermatum. Thus, the biocontrol activities of bacterial strains against target diseases could be expressed consistently and effectively through the enhancement or maintenance of the populations of the antagonistic strains. Accordingly, the tested strains in this study may need to preserve certain population levels in the pepper rhizosphere or root to consistently elicit ISR against C. acutatum infection. Therefore, to achieve consistent biocontrol efficacy against anthracnose and Phytophthora blight of pepper in fields or plastic houses, which exhibit dynamic environmental changes, careful evaluation of the temporal population changes of the treated strains in pepper rhizosphere or root is needed.
In this study, three of four selected strains (YJR27, YJR92, and YJR102) increased pepper fruit numbers after inoculation in the field tests, and these strains increased red fruit weights at the first harvest in the plastic house tests. The increased fruit yield might be dependent on disease reduction by the antagonistic strains, because yield reduction was not observed before inoculation. Moreover, the plastic house tests revealed that the strains exhibited plant growth-promoting ability in terms of pepper fruit yield. It is known that plant growth promotion can be achieved directly by enhancing the utilization of nutrients and minerals, or by regulating plant hormones including indole-3-acetic acid (IAA), cytokinins, and ethylene or indirectly by disease suppression via biocontrol agents (Gray and Smith, 2005; Mia et al., 2010; Shanmugam et al., 2011; Zheng et al., 2011). Thus, these results show that the strains YJR27, YJR92, and YJR102 have the ability to improve plant growth and health by protecting against the tested plant diseases in this study.
When these selected strains were identified using FAME and 16S rRNA gene sequence analyses, strain YJR27 was determined as P. otitidis, strain YJR92 as P. putida, strain YJR102 as T. tyrosinosolvens and strain YJR107 as N. capsulatum. It is well known that many strains belonging to the genus Pseudomonas have biological traits such as antibiotic production, competitive colonization, ISR, and plant growth promotion (Weller, 2007). Within the genus Pseudomonas, P. putida has been reported as an effective biocontrol bacterial species against various plant diseases including Rhizoctonia and Pythium root rot of wheat (Mavrodi et al., 2012). Likewise, P. otitidis has been found in various environments including water waste, and can produce N-acylhomoserine lactones (AHLs), IAA, and bacteriocin (Mehri et al., 2011). However, T. tyrosinosolvens, which belongs to Actinomycetes, has not previously been reported as an antagonistic bacterial species against plant diseases; however, a strain of species belonging to the genus Tsukamurella has been isolated from the gut of Japanese honeybee (Apis cerana japonica) (Yoshiyama and Kumura, 2009), and from the samples of patients with cardiac pacemaker implants and chronic lung infections (Yassin et al., 1997). In the case of N. capsulatum, Halfeld-Vieira et al. (2008) observed that the species was capable of reducing late blight of tomato caused by Phytophthora infestans under field condition. The results of bacterial identification of the selected strains, therefore, indicate that the use of the sequential screening procedure has resulted in the selection of diverse bacterial resources for controlling pepper diseases and promoting plant growth.
Taken together, these results indicate that the sequential screening procedure is a rapid and reliable method to select effective antagonistic bacterial strains against P. capsici in pepper plants. This screening procedure allowed us to obtain four effective biocontrol strains (P. otitidis YJR27, P. putida YJR92, T. tyrosinosolvens YJR102, and N. capsulatum YJR107) against Phytophthora blight of pepper. These strains also had biocontrol activities against anthracnose, and the strains YJR27, YJR92, and YJR102 also showed plant growth-promoting activities on pepper fruit. Thus, these bacterial strains may be promising biocontrol agents against Phytophthora blight and anthracnose of pepper.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






