Application of Arbuscular Mycorrhizal Fungi during the Acclimatization of Alpinia purpurata to Induce Tolerance to Meloidogyne arenaria
Article information
Abstract
An experiment was conducted to evaluate the tolerance of micropropagated and mycorrhized alpinia plants to the parasite Meloidogyne arenaria. The experimental design was completely randomized with a factorial arrangement of four inoculation treatments with arbuscular mycorrhizal fungi (AMF) (Gigaspora albida, Claroideoglomus etunicatum, Acaulospora longula, and a non-inoculated control) in the presence or absence of M. arenaria with five replicates. The following characteristics were evaluated after 270 days of mycorrhization and 170 days of M. arenaria inoculation: height, number of leaves and tillers, fresh mass of aerial and subterranean parts, dry mass of aerial parts, foliar area, nutritional content, mycorrhizal colonization, AMF sporulation, and the number of galls, egg masses, and eggs. The results indicated a significant interaction between the treatments for AMF spore density, total mycorrhizal colonization, and nutrient content (Zn, Na, and N), while the remaining parameters were influenced by either AMF or nematodes. Plants inoculated with A. longula or C. etunicatum exhibited greater growth than the control. Lower N content was observed in plants inoculated with AMF, while Zn and Na were found in larger quantities in plants inoculated with C. etunicatum. Fewer galls were observed on mycorrhized plants, and egg mass production and the number of eggs were lower in plants inoculated with G. albida. Plants inoculated with A. longula showed a higher percentage of total mycorrhizal colonization in the presence of the nematode. Therefore, the association of micropropagated alpinia plants and A. longula enhanced tolerance to parasitism by M. arenaria.
Introduction
Tropical ornamental plants exhibit exotic forms, durability, and high productivity, which have contributed to the expansion of the market (Lamas, 2002). Pernambuco is one of the largest Brazilian producers of tropical flowers (Ferreira et al., 2002). Among these, Alpinia purpurata (Viell.) Schum (alpinia or panama) has stood out with regard to production and commercialization. However, on many plantations the plants are parasitized by Meloidogyne phytonematodes that cause growth reduction, thus resulting in rachitic plants and loss of productivity (Bala and Hosein, 1996).
Micropropagation is a plant production method that allows the attainment of large numbers of highly productive, genetically homogeneous, and pathogen-free individual plants in a short period of time and with limited physical space (Grattapaglia and Machado, 1998). The formation of mycorrhizae improves the nutritional condition of the plant, and makes it more resistant to biotic and abiotic stresses (Azcón-Aguilar and Barea, 1997a). In tropical regions, arbuscular mycorrhizae, which are formed by arbuscular mycorrhizal fungi (AMF), are most commonly found in native plants and plants of economic interest. Plants benefit from AMF mycorrhization during the seedling stage as well as during the acclimatization of micropropagated plants via various culturing methods (Borkowska, 2002; Declerck et al., 2002).
Diverse members of the soil biota associated with plants interact among themselves and with other plants, thus influencing plant fitness. Interactions among AMF, plants, and nematodes are complex, and the nature and effects of these interactions depend on the species (Castillo et al., 2006; Hol and Cook, 2005) and the environmental conditions involved (Calvet et al., 2001). The effects of AMF on the growth of plants infected with Meloidogyne nematodes vary, but are generally positive (Borowicz, 2001; Campos et al., 2013; Maia et al., 2006).
The production of micropropagated seedlings of AMF-inoculated ornamental tropical plants may improve the control of Meloidogyne spp. that parasitize the plants. The objective of this study was to evaluate the tolerance of micropropagated and mycorrhized A. purpurata seedlings to Meloidogyne arenaria (Neal.) Chitwood.
Materials and Methods
Substrate
Soil (yellow latosoil), disinfected with Bromex ® (98% methyl bromide and 2% chloropicrin), with the following characteristics was used 82% sand, 13% silt, and 5% clay; pH 7.1; 21 mg dm−3 P; 1.1 cmolc dm−3 K; 4.7 cmolc dm−3 Ca; 1.3 cmolc dm−3 Mg; 0.07 cmolc dm−3 Na; 13.3 g dm−3 MO (organic material), and 9.64 cmolc dm−3 CTC (cationic exchange capacity).
Vegetable material
Micropropagated A. purpurata plantlets were multiplied in MS medium (Murashige and Skoog, 1962) supplemented with 4 mg l−1 BAP (6-benzylaminopurine) at the Biotechnological Laboratory of Embrapa Semiarid (Brazilian Agricultural Research Corporation; https://www.embrapa.br/semiarido).
AMF
Gigaspora albida N. C. Schenck & G. S. Sm. (UFPE 01), Claroideoglomus etunicatum W. N. Becker & Gerd. (UFPE 06), and Acaulospora longula Spain & N. C. Schenck (UFPE 21), stored at the AMF Inoculants Bank of the UFPE (Federal University of Pernambuco), were used. The inoculants were multiplied in vases with sand and vermiculite (1:1 v/v), irrigated with Hoagland nutritive solution (Jarstfer and Sylvia, 1992), and supplemented with Tris-HCl (pH 6.5), and Panicum miliaceum L. was used as a host. After inoculum multiplication, the number of AMF infective propagules was estimated using the method of Feldmann and Idczak (1994).
Inoculation with AMF
At the start of acclimatization, the plants were transplanted from in vitro conditions to pots containing 2 kg soil plus 1,500 infective AMF propagules. After 125 days, the plants were transferred to vases with 3 kg soil capacity.
Nematode attainment
To obtain the Meloidogyne suspension, infected A. purpurata root samples were collected at Embrapa-SNT (Embrapa Technology Transference) in Petrolina, Pernambuco, Brazil. The samples were carefully washed with tap water, and the eggs were extracted using the method of Hussey and Barker (1973). Eggs were counted under a stereomicroscope to determine the number of eggs per microliter.
Nematode inoculation
After three months of acclimatization, a watery suspension with approximately 3,000 M. arenaria eggs and juveniles were introduced to the root region of A. purpurata plants.
Experimental conditions
The experiment was conducted at the greenhouse at Embrapa Semiarid, Petrolina, Pernambuco, Brazil, under controlled environmental conditions (temperature, 27 ± 2°C; relative humidity, 75%; luminosity, 250–560 μmol m−2 s−1).
Evaluation
The following characteristics were evaluated after 270 days of AMF inoculation: height, number of leaves, number of tillers, fresh and dry mass of aerial parts and the fresh subterranean mass, foliar area, mycorrhizal colonization, AMF spore density, and the number of egg masses, galls and nematode eggs. The foliar area was measured with a Li 3100 apparatus (Li-Cor Inc., Lincoln, NE, USA). Dry biomass was obtained after oven drying (65°C) until a constant weight was obtained. This material was sent to the Soil and Plant Analysis Laboratory of Embrapa Semiarid to evaluate the nutritional content of the aerial parts (N, P, K, Ca, Mg, B, Cu, Fe, Mn, Zn, and Na). The extent of hyphae, arbuscular, vesicular, and total mycorrhizal colonization was estimated using the method of McGonigle et al. (1990) with diaphanized roots in 10% KOH, which were stained with 0.03% Chlorazol Black (Brundrett et al., 1984). AMF spores were extracted from the soil by decantation and wet sieving, followed by centrifugation in water and saccharose (Gerdemann, 1963; Jenkins, 1964). The specimens were then quantified under a stereomicroscope (40×). The number of galls and the egg mass quantity were evaluated by direct counting with the naked eye, and the number of extracted eggs from the roots was determined with a stereomicroscope (Hussey and Barker, 1973).
Experimental design
The experimental design was completely randomized with a factorial arrangement of four AMF inoculation treatments (non-inoculated control or inoculation with A. longula, G. albida, or C. etunicatum) and two treatments with nematodes (presence [CN] or absence [SN] of M. arenaria). Five replicates were conducted, totaling 40 experimental units.
Statistical analysis
The data were subjected to variance analyses using the Statistica program (Statsoft, 1997), and the averages were compared using Tukey’s test with 5% probability. The values (x), in percentages and in numbers, were transformed into arcsin x/100 and log x + 1 values, respectively.
Results and Discussion
Growth parameters
The height and number of tillers only differed significantly in treatments with nematodes (Table 1). However, the fresh mass of the aerial and subterranean parts, the dry mass of aerial parts, and the foliar areas differed in AMF and nematode treatments separately (Table 1, 2). The height, fresh mass of aerial and subterranean parts, dry mass of aerial parts, and foliar areas of alpinia plants were higher in plants that were not infected by nematodes (Table 1). Conflicting results were observed in plants with respect to the number of tillers, which exhibited larger quantities in seedlings with nematodes (Table 1). The presence of Meloidogyne in alpinia seems to have been stimulated by tillering. This behavior likely occurred as a reaction of plants to nematode infection, and new ramifications (tillers) were emitted to secure survival in the presence of Meloidogyne. Dinardo-Miranda et al. (2001) observed large numbers of tillers in control plants without the addition of herbicide and nematicide in sugarcane plantations (Saccharum officinarum L. var. RB845257) infested with Meloidogyne javanica (Treub.) Chitwood.
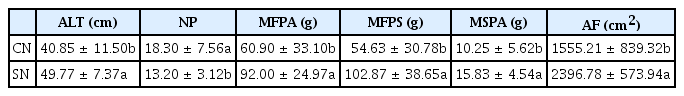
Growth variables of Alpinia, in the presence (CN) or absence (SN) of Meloidogyne arenaria, independent of arbuscular mycorrhizal fungi inoculation, after 270 days in the greenhouse
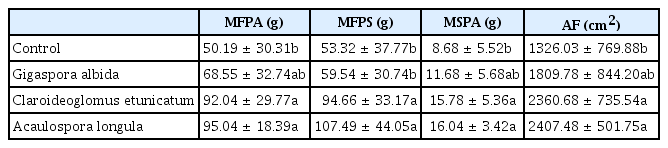
Growth variables of non-inoculated or arbuscular mycorrhizal fungi-inoculated alpinia plants, independent of the presence of Meloidogyne arenaria, after 270 days in the greenhouse
Plants inoculated with A. longula or with C. etunicatum showed greater growth than the control plants (Table 2), and this resulted in higher fresh mass of aerial and subterranean parts and the dry mass of aerial parts and foliar areas in alpinia plants inoculated with these fungi than was observed following the other treatments. No mycorrhizal benefits were observed in the acclimatization experiment (100 days), which used the same soil and AMF isolates (Silva et al., 2006). However, in this experiment (evaluated after 270 days), inoculation with A. longula and C. etunicatum benefited the host plant to a greater extent than did the non-inoculated treatment. Estaún et al. (1999) observed that the benefits of AMF application in micropropagated peach trees (Prunus persica (L.) Batsch. × Prunus amygdalus Stokes.) were exhibited after only 14 months of inoculation, suggesting that plant growth promotion may occur at a later stage.
Nutrient content
There were significant interactions between the treatments regarding N, Na, and Zn content in aerial alpinia parts (Table 3). The Cu and Mg quantities in the aerial parts differed in AMF (Table 4) or nematode (Fig. 1) treatments separately, and B, Mn, and P content in the aerial parts was only affected by the AMF treatment (Table 4). The content of Mg and Cu was higher in alpinia plants that were not infected by nematodes (Fig. 1).
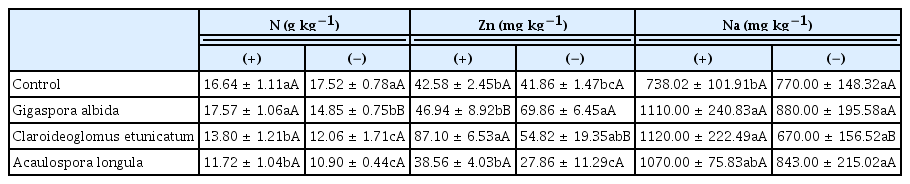
Nutritional content of non-inoculated or arbuscular mycorrhizal fungi-inoculated aerial parts of alpinia in the presence (+) or absence (−) of Meloidogyne arenaria after 270 days in the greenhouse

Nutritional content of the aerial parts of alpinia, with or without arbuscular mycorrhizal fungi (control), independent of inoculation with Meloidogyne arenaria, after 270 days in the greenhouse
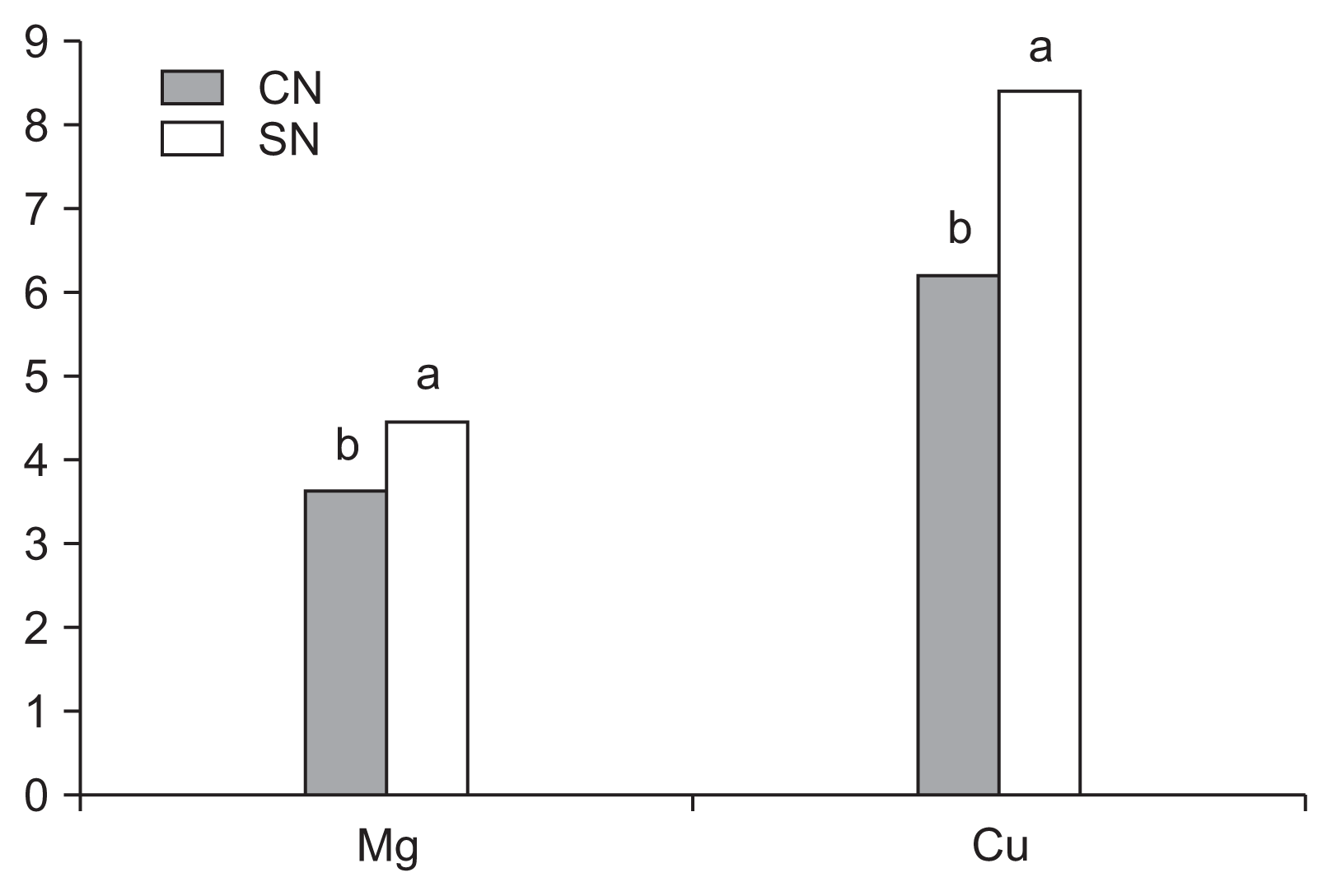
Content of Mg (g kg−1) and Cu (mg kg−1) in alpinia, with (presence, CN) or without nematodes (absence, SN), independent of arbuscular mycorrhizal fungi inoculation, after 270 days in the greenhouse. Averages followed by the same letter do not differ significantly based on Tukey’s test at the 5% significance level.
The results obtained by different authors regarding the benefits of mycorrhization in plants exposed to Meloidogyne infections are diverse and depend on the type of AMF inoculant. In Cucumis sativus L. ‘Zhongnong 16’ infected with Meloidogyne incognita, plants inoculated with Glomus versiforme (P. Karsten) S.M. Berch showed an increase in biomass as well as in increase in the absorption of P, Fe, and Cu, which did not occur in Glomus intraradices Schenck & Smith (Zhang et al., 2008) treatments. In the present experiment, C. etunicatum and A. longula caused greater growth. However, G. albida was less effective, and did not increase plant growth in the presence of M. arenaria. This result supported the hypothesis that the tolerance of mycorrhized plants to phytonematode parasitism is dependent on the AMF species applied (Azcón-Aguilar and Barea, 1997b).
The content of K, Fe, and Ca in the aerial parts of alpinia plants did not differ in the treatments. Neither AMF nor nematodes appeared to influence the content of these nutrients in the aerial parts of alpinia plants. Carneiro et al. (2002) observed that the quantity of Ca in the aerial parts of soy (Glycine max (L.) Merr.) was not affected by the presence of M. incognita or M. javanica, and Jaizme-Vega et al. (1997) obtained similar results in mycorrhized banana trees infected with M. incognita.
The quantity of N was lower in plants with Meloidogyne that were inoculated with C. etunicatum or A. longula, while all AMF treatments resulted in smaller N quantities in non-Meloidogyne-infected plants than in the control. In the G. albida treatment, the N quantity was higher in plants with nematodes than those without nematodes (Table 3). This smaller N quantity in the aerial parts might be related to the dilution effect that occurs in plants with AMF (Al-Karaki, 2006). In addition, plants infected with Meloidogyne absorbed more N, and translocation to the aerial parts was normally not affected (Carneiro et al., 2002).
A large quantity of Zn was observed in plants infected with Meloidogyne and mycorrhized with C. etunicatum, and this differed from the effects observed for the other treatments. However, in plants without Meloidogyne, a large quantity of Zn was found in the G. albida treatment in comparison to the control and the A. longula treatment (Table 3). In plants inoculated with G. albida, a high Zn content was registered in plants without nematodes than in those with nematodes, but plants inoculated with C. etunicatum exhibited the inverse behavior (Table 3). Generally, plants inoculated with AMF, even in the presence of Meloidogyne, showed a high Zn content, but it depended on the AMF isolate used (Zhang et al., 2008).
In the presence of Meloidogyne, the Na content was higher in plants inoculated with G. albida or C. etunicatum than in the control (Table 3). A lower quantity of Na was documented in plants inoculated with C. etunicatum without Meloidogyne than in infected plants (Table 3). This nematode appears to stimulate the absorption and translocation of Zn and Na, especially when plants are inoculated with C. etunicatum, because high Zn and Na content was observed in aerial Alpinia plants in the presence of the two organisms.
Large quantities of Mg and Cu were observed in the aerial parts of alpinia plants associated with A. longula, and large quantities of Mn were detected in plants inoculated with C. etunicatum. However, B and P content was lower in the A. longula and C. etunicatum treatments, respectively (Table 4). The P content in the aerial parts of plants inoculated with AMF was generally higher than that of the control (Jaizme-Vega et al., 1997), and this was the case with other nutrients such as Cu, Mg, and Mn (Zhang et al., 2008) that were dependent on the AMF isolate used. However, in the current study, the plants inoculated with C. etunicatum showed a lower quantity of P than those inoculated with other AMF isolates. This may have resulted from lower P absorption or translocation for the aerial parts of plants inoculated with C. etunicatum. Some micronutrients were also found in lower quantities in plants with AMF, and this may be related to the absorption or translocation process (Carneiro et al., 2002). For instance, some of these nutrients may have exhibited normal absorption, but translocation to the aerial parts was affected, thus resulting in lower content values.
AMF parameters
There was significant interaction between the treatments regarding AMF spore density and total mycorrhizal colonization, while hyphal and arbuscular mycorrhizal colonization were only affected by the AMF treatment (Table 5). In plants without nematodes, all of the AMF isolates produced more spores than the control. However, in the pathogen treatments, only C. etunicatum and A. longula exhibited greater sporulation, and this differed from the control and the G. albida treatments (Table 5). Moreover, the presence of the nematode induced a reduction in the quantity of spores produced by A. longula and by G. albida (Table 5). In the rhizosphere of sweet passion fruit plants, there was also a reduction in sporulation of Scutellospora heterogama (Nicol. & Gerd.) Walker & Sanders in the presence of M. incognita (Anjos et al., 2010). Cofcewicz et al. (2001) observed greater sporulation of Gigaspora margarita than C. etunicatum in tomato plants infected with M. javanica, which is contrary to the results of the current study, in which C. etunicatum produced more spores than G. albida. The amplitude of the responses to the interaction between nematodes and AMF, other than intra- and interspecific variance, depends on the plant species and the soil used.
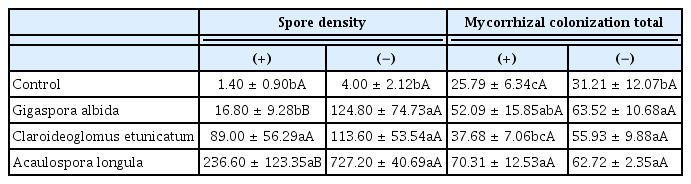
Arbuscular mycorrhizal fungi (AMF) spore density (50 g−1 of soil) and mycorrhizal colonization of vesicles and total colonization (%) in the rhizosphere of alpinia plants (non-inoculated or AMF-inoculated) in the presence (+) or absence (−) of Meloidogyne arenaria after 270 days in the greenhouse
The percentage of mycorrhizal colonization by vesicles did not differ between the treatments (Table 5). However, the colonization by hyphae or arbuscular fungi only differed in AMF treatments (Fig. 2). All AMF treatments exhibited greater mycorrhizal colonization by hyphae than the control, and higher values were observed in plants associated with G. albida and A. longula. However, contrasting results were observed for arbuscular colonization.
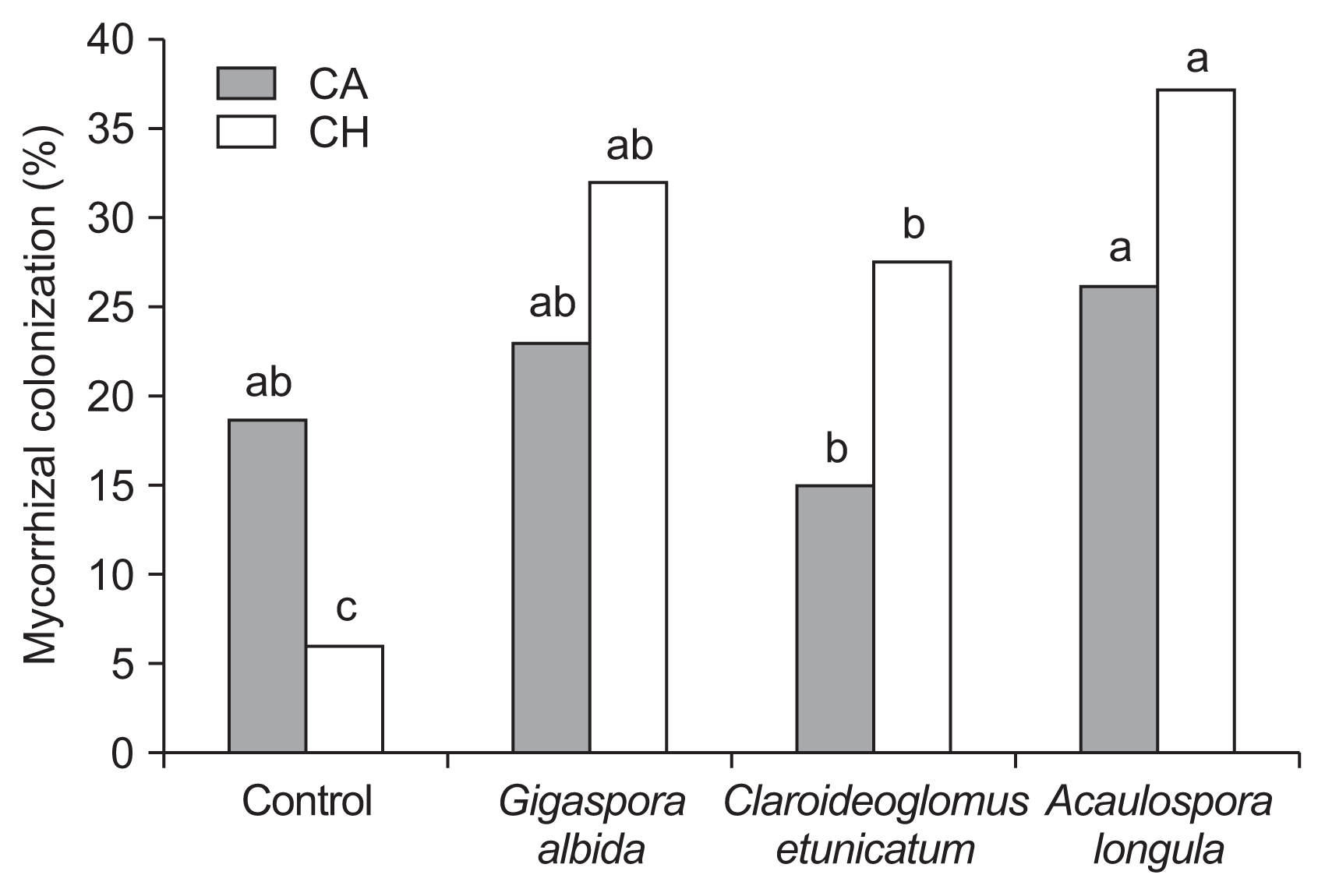
Mycorrhizal colonization of arbuscules (CA) and hyphae (CH) (%) in the roots of alpinia plants (non-inoculated or arbuscular mycorrhizal fungi-inoculated), independent of inoculation with nematodes, after 270 days in the greenhouse. Averages followed by the same letter do not differ significantly based on Tukey’s test at the 5% significance level.
Nevertheless, total colonization showed distinct interactions (Table 5). For instance, in the presence of nematodes, plants inoculated with A. longula showed a higher total mycorrhizal colonization percentage, and this differed from plants inoculated with C. etunicatum and from the control. In the absence of the pathogen, all AMF treatments differed from the control. Zhang et al. (2008) observed a larger percentage of mycorrhizal colonization in C. sativus inoculated with G. intraradices than in treatments with Glomus mosseae and G. versiforme when in the presence of M. incognita. However, in the absence of the pathogen, greater colonization was observed in the roots of plants inoculated with G. intraradices or with G. mosseae than those inoculated with G. versiforme.
Meloidogyne parameters
Mycorrhized plants showed lower numbers of galls per gram of root than the control treatment (Table 6). However, the production of egg masses and eggs per gram of root by phytonematodes was less for plants inoculated with G. albida than the control treatment (Table 6). In tomato plants (Lycopersicon esculentum L. ‘Tounvi’) infected with Meloidogyne spp., a reduction in the quantity of galls and an increase in the number of eggs in the treatments with G. mosseae or Acaulospora spinosa C. Walker & Trappe were observed (Affokpon et al., 2011). The present study showed a similar result, and lower numbers of galls were found in all AMF treatments. However, in treatments with C. etunicatum and A. longula, the quantity of eggs was higher.

Number of galls (NG), egg masses (NMO), and eggs (NO) in the roots of Alpinia purpurata plants, non-inoculated (control) or arbuscular mycorrhizal fungi-inoculated, infected with Meloidogyne arenaria after 270 days in the greenhouse
A reduction in gall and egg production by M. incognita was observed in banana trees in the presence of G. mosseae (Jaizme-Vega et al., 1997), in tomato plants associated with Glomus coronatum Giovannetti (Diedhiou et al., 2003), and in peach trees inoculated with G. mosseae, G. intraradices, or C. etunicatum (Calvet et al., 2001). In general, a reduction in the reproduction of the phytonematodes was observed. However, the presence of AMF could increase the quantity of galls and the number of eggs, and this was reported in the roots of tomato plants infected with M. javanica (Cofcewicz et al., 2001) and in the roots of peanuts (Arachis hypogaea L.) infected with M. arenaria (Carling et al., 1995). Furthermore, the AMF isolates used should also be considered, since their effects on gall production and nematode reproduction differ (Affokpon et al., 2011).
In the present study, fewer galls were present in plants inoculated with AMF, while the quantities of egg masses and eggs were significantly lower in the G. albida treatment. However, a tendency to reduce these parameters in the A. longula treatment was observed. On the other hand, treatment with C. etunicatum, relative to these parameters was very similar to the non-treatment control. Plants inoculated with C. etunicatum showed similar growth to those inoculated with A. longula, and greater growth was observed than for plants from the control treatment and those inoculated with G. albida. These outcomes might have resulted from better nutrition that allowed greater nematode reproduction, because the nutrients could be allocated by the nematode for its development and reproduction. Therefore, plants that have better nutrition could stimulate nematode development (Hussey and Roncadori, 1982).
Final considerations
The increase in vegetable growth in the presence of Meloidogyne, promoted by A. longula inoculation, may be related to high radicular colonization. Namely, the occupation of a large area of the radicular cortex could diminish the physical space for radicular infection and, consequently, the production of galls. Decreased available radicular space and increased nutrient input (higher vigor) were suggested by Lindermann (2000) as mechanisms related to the mitigation of negative nematode effects by AMF. On the other hand, the lower radicular colonization by C. etunicatum in alpinia could lead to the opposite effect observed in A. longula, resulting in a lower capacity to block the action of the nematode and the production of a larger number of galls, as was observed in roots mycorrhized by C. etunicatum.
Habte et al. (1999) considered AMF species with the capacity to induce plant resistance to nematodes ideal, namely, reduced nematode reproduction and concurrent host growth production. However, occurrences of AMF-induced resistance are less common than those of tolerance, in which both the nematode and plant develop well. In the current study, plants inoculated with A. longula or C. etunicatum grew well in concert with the nematode. Therefore, these species have the capacity to induce tolerance to M. arenaria, which can be used as an alternative method to diminish the damage caused by this pathogen.
Acknowledgments
The authors would like to thank CAPES for PhD scholarships to M.A.S. Campos and F.S.B da Silva, and CNPq for financial support and a scholarship awarded to L. C. Maia. We would also like to thank the Laboratório de Biotecnologia e Microbiologia of Embrapa Semiarid for logistic support for study development, and Dr. Regina M. D. G. Carneiro of Embrapa Recursos Genéticos e Biotecnologia for nematode identification.