Morphometric Variation in Pine Wood Nematodes, Bursaphelenchus xylophilus and B. mucronatus, Isolated from Multiple Locations in South Korea
Article information
Abstract
Intraspecific variation in morphometry of pine wood nematodes Bursaphelenchus xylophilus and B. mucronatus in relation to geographical locations in South Korea was investigated using morphometric characters (body length, a, b and c ratio, stylet length, and spicule length for a male nematode and V (%) value for a female nematode). B. xylophilus was isolated from Pinus thunbergii in Jinju (1998), Ulsan (2000), Yangsan (2000), Mokpo (2001) and Jeju (2004), and from P. densiflora in Gumi (2001). B. mucronatus was isolated from P. thunbergii in Jinju (1991) and from P. densiflora in Milyang (2001). The body length of male and female B. xylophilus had the highest coefficient of variability and showed significant differences among geographical locations. The V (%) value for female B. xylophilus showed the lowest coefficient of variability, changing little with geographical area and host plant. All morphometric characters in B. mucronatus except for stylet length and female body length showed no significant differences between locations or hosts, suggesting they may not be affected by geographical area or host plant.
Outbreaks of pine wilt disease in East Asian countries are caused by the exotic pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle, which was previously endemic to North America. In South Korea, pine wilt disease was first discovered in a pine forest in Busan city in 1988, following introduction of this nematode, and has since spread across the country. The spread of the nematode has been facilitated by the vector Monochamus alternatus Hope in the southern part of South Korea (Yi et al., 1989), and by M. saltuarius Gebler in northern part of South Korea (Kim et al., 2007). Following its initial discovery in South Korea, B. xylophilus has since been detected in dead pine trees in the locations of Jinju (1998), Ulsan (2000), Yangsan (2000), Mokpo (2001), Gumi (2001), Jeju (2004), Seoul (2007), Yeosu (2010), and by 2011 it had spread to 44 other locations (Choi et al., 2012). This pathogenic pine wood nematode has also been introduced to the European Union via numerous sources, including round wood, packaging, and wood chips imported from infected areas (Evans et al., 1996). Braasch (2001) analyzed five morphometric characters of the genus Bursaphelenchus collected in the European Union, and showed that B. xylophilus and B. mucronatus Mamiya and Enda exhibited a wide variation in morphometric characters such as body length. In China, Bursaphelenchus spp. including B. xylophilus were detected from imported wooden packaging in Ningbo Port (Gu et al., 2008). Yin et al. (1988) and Yusheng et al. (2002) reported that some essential morphometric characters of the genus Bursaphelenchus collected in Hunan and Guangdong province in China, respectively.
Morphometric characters of the plant pathogenic nematode Pratylenchus penetrans Filip. and Schuurm-Stekh, such as body length, body width, esophagus length, stylet length, and a ratio, can vary according to a variety of environmental factors, including host plant (Tarte and Mai, 1976). Townshend (1991) reported that the morphological characteristics of P. penetrans were affected by the host to a greater extent than by geographical area. Of those variable morphometric characters, the V (%) value of P. penetrans was insufficiently affected by host plant for it to be of diagnostic value (Tarte and Mai, 1976). Tarjan and Frederick (1978) reported that the stylet lengths of P. brachyurus and P. coffeae had the lowest variability of the characters studied. In the case of Meloidogyne arenaria (Neal) Chitwood, the values of most morphometric characters were significantly different between populations (Cliff and Hirschmann, 1985). Here, we investigated the intraspecific variation of morphometric characters of B. xylophilus and B. mucronatus from different geographical locations and host plants to establish the extent of morphological variation in populations of these two species in South Korea.
Materials and Methods
The pine wood nematode, B. xylophilus, was Pine wood nematodes, B. xylophilus, were isolated from dead pine trees (Pinus thunbergii Parl.), infected with pine wilt disease, in Jinju, Ulsan, Yangsan, Mokpo, and Jeju, and from P. densiflora Sieb. et Zucc. in Gumi. B. mucronatus was similarly isolated from P. thunbergii in Jinju and from P. densiflora in Milyang, South Korea (Fig. 1). Several adult nematodes were randomly collected from various stages of nematodes which were extracted from several dead trees at each location. All collected pine wood nematodes were fixed with F.G. 4:1 solution (80% formalin and 20% glycerin), which was heated to 80 °C, and then transferred to anhydrous glycerin to prepare permanent specimens using modified Seinhorst’s rapid glycerin method (Seinhorst, 1959). Body length, maximal body diameter, distance from anterior end to pharynx-intestine junction, tail length, stylet length, spicule length and distance from anterior end to vulva of each pine wood nematode were measured. Morphometric character of a ratio, b ratio and c ratio were calculated from body length divided by maximal body diameter, by distance from anterior end to pharynx-intestine junction and by tail length, respectively. V (%) value was also calculated from a percentage of distance from anterior end to vulva divided by body length.
The locations where pine wood nematodes were collected in South Korea. Bursaphelenchus xylophilus was collected from Jinju, Ulsan, Yangsan, Mokpo, Gumi, and Jeju. East Asian types of B. mucronatus was collected from Jinju and Milyang.
For each gender of B. xylophilus, the morphometric characters of B. xylophilus were separately compared among the six locational groups using ANOVA and Tukey’s honestly significant difference method. The morphometric characters of B. mucronatus were compared between locational groups using a 2-tailed t-test (P < 0.05).
Results and Discussion
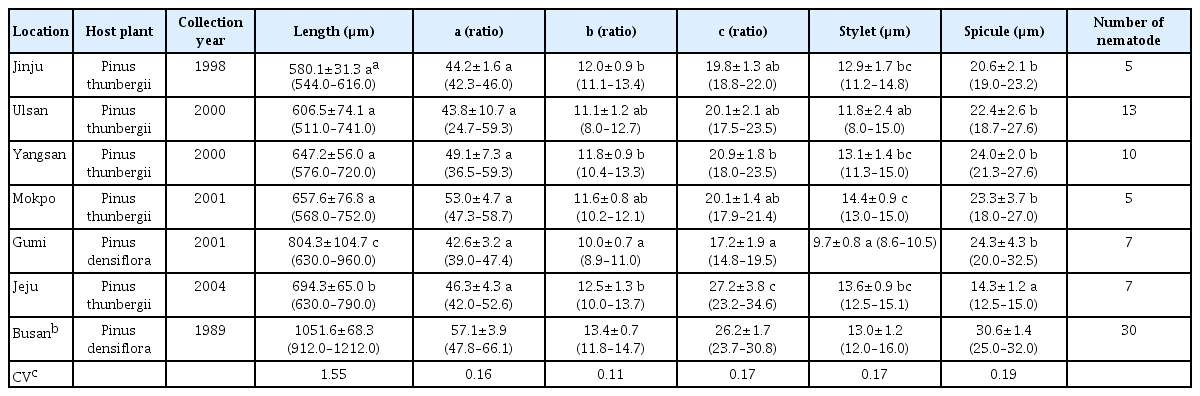
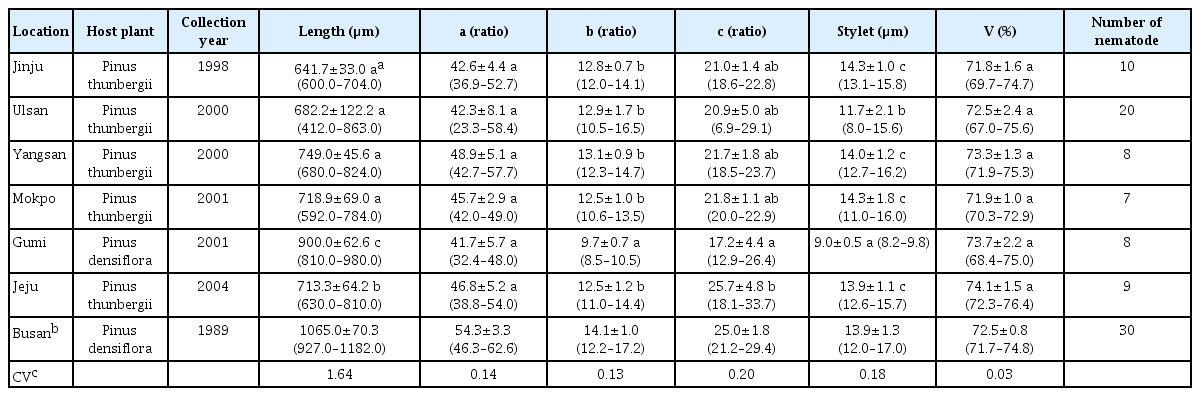
The morphometric features of male and female B. xylophilus in each locational area are shown in Tables 1 and 2, respectively. The mean body length of male B. xylophilus from the six locations was divided into three subgroups: Gumi, Jeju, and the other four locations. The mean body length of male B. xylophilus from the Gumi location was the longest among the six locational groups, with the Jeju location nematodes being the second longest. In the other four locations (Jinju, Ulsan, Yangsan, and Mokpo), the mean body length of male B. xylophilus did not significantly differ from each other. The mean body length of female B. xylophilus was divided into the same three subgroups as for males. The mean body length of the six locational groups were smaller than those from the Busan location in both males and females (Choi and Moon, 1989), but overlapped with the mean body length of male and female B. xylophilus in Japan that were 730 μm and 810 μm, respectively (Mamiya and Kiyohara, 1972). The mean body length also overlapped with those of nematodes found in Japanese packaging wood in China, which were 870 μm and 1021 μm, respectively (Li et al., 2009). Our data falls within the range of male and female B. xylophilus body lengths that were 590 – 1300 μm and 447 – 1290 μm, respectively, in the report of Braasch (2001). There was wide variation of B. xylophilus body length. In case of P. penetrans, wide variation of body length can be explained by host plants or the geographical origin of the nematode populations (Tarte and Mai, 1976; Townshend, 1991). A noteworthy feature of the Gumi locational group is that both male and female B. xylophilus had the longest body length and were isolated from P. densiflora, whereas the other nematodes were isolated from P. thunbergii. Tarte and Mai (1976) showed that nematode body length was influenced by host plants, thus the body length of B. xylophilus in Gumi may be related to its host plant. A unique aspect of the Jeju locational group is that Jeju is an island found along the Korean Peninsula (Fig. 1), and this feature might be related to the fact that the body lengths of both male and female B. xylophilus in the Jeju location were the second longest among the nematodes we analyzed. The coefficient of variability for body length was 1.55 and this was noticeably the largest among the six morphometric characters measured. This suggests that B. xylophilus body length is the most variable morphometric character in this study, despite the fact that the four locational groups of Jinju, Ulsan, Yangsan and Mokpo were not significantly differentiated from one another. The geographical variation of pine wood nematode body length, however, seems not to be related to genetic variation as the nematode population in Ulsan groups in the same subclade of Busan, rather than that in Jinju, which is located in a separate clade (Jung et al., 2010).
Morphometric data of male Bursaphelenchus xylophilus collected in South Korea. Mean ± SD are shown with the range of measurement in parentheses
Morphometric data of female Bursaphelenchus xylophilus collected in South Korea. Mean ± SD are shown with the range of measurement in parentheses
The mean a ratios of male and female B. xylophilus across the six locations were not significantly different from each other. The a ratio of B. xylophilus in this study was among the least variable morphometric characters. In the study of Li et al. (2009), however, the a ratio of three isolates of female B. xylophilus from Japan, Canada and China was a useful character for delineating populations. It can be supposed that the geographic distances between six locational areas in South Korea were not sufficiently substantial to result in locational differentiation of the a ratio.
The mean b ratio of male B. xylophilus were not sufficiently variable to identify locational groups in this study. The mean b ratio of female B. xylophilus from the Gumi location was significantly smaller than those of other five locational groups, and the host plant of B. xylophilus from the Gumi location was P. densiflora, whereas those from other five locational groups were P. thunbergii. Based on this result, the mean b ratio of female B. xylophilus seems to have relation to a species of host plants.
The mean c ratio of male B. xylophilus from the Jeju location was significantly larger than that of other five locational groups. Based on this result, the mean c ratio of male B. xylophilus seems to have relation to locational area. The mean c ratio of female B. xylophilus, however, was not sufficiently variable to identify locational groups in this study, whereas Li et al. (2009) found it served as another useful character for separating populations.
Stylet length of B. xylophilus is known as a useful character for population identification (Li et al., 2009), while that of some Pratylenchus spp. is known as the least variable character for species identification (Roman and Hirschmann, 1969; Tarjan and Frederick, 1978). In this study mean stylet lengths of male B. xylophilus did not separate the locational groups. In contrast, the mean stylet lengths of female separated significantly the locational groups into three subgroups: Gumi, Ulsan, and the other four locations.
The mean spicule length of male B. xylophilus of the Jeju locational group was significantly shorter than that of other five locational groups. Accordingly, this morphometric character has been used for population identification (Li et al., 2009). V (%) values of female B. xylophilus across the six locational groups were not significantly different from each other, and have the lowest coefficient of variability among the six morphometric characters analyzed in this study. These results suggest that V (%) value is minimally affected by geographical locations and host plants. A similar pattern was observed in six species of Pratylenchus, where V (%) value had the lowest coefficient of variability (Roman and Hirschmann, 1969). In addition, Tarte and Mai (1976) found that V (%) value of P. penetrans was not sufficiently affected by host plants to be of diagnostic value.
The morphometric features of body length, b and c ratio, stylet length, and spicule length of male B. xylophilus and those of body length, b and c ratio, and stylet length of female B. xylophilus isolated from Jeju were significantly different from those of Gumi. These differences might be induced (or maintained) by the sea barrier as Jeju is an isolated island, and may also be due to differences in host plants.
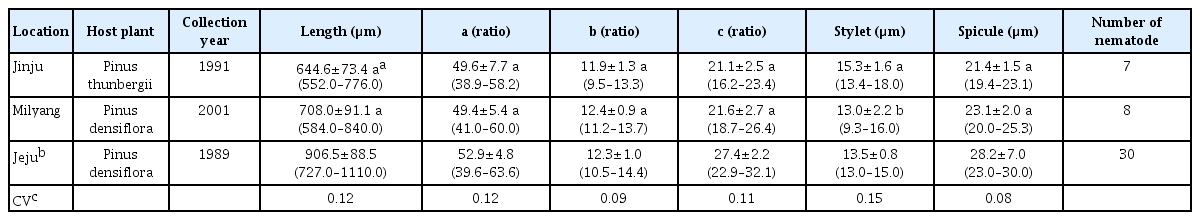
In the case of B. mucronatus, male stylet lengths and female body lengths showed significant differences between the Jinju and Milyang locational groups (Tables 3 and 4). The mean body length of the Jinju and Milyang locational groups were smaller than those from Jeju in both males and females (Choi and Moon, 1989), but overlapped with those of B. mucronatus in Japan (Mamiya and Enda, 1979). The ratios of a, b and c in males and females, the spicule length of males, and the V (%) value for females of B. mucronatus did not differ between the two locational groups, suggesting that these characters are minimally affected by geographic locations or host plants. Based on the study of Li et al. (2009), the three populations of both male and female B. mucronatus (East-Asian type) were also not separated by five different morphometric characters. The V (%) value had the lowest coefficient of variability among the six morphometric characters, suggesting it might be useful for species identification. However, there was no significant difference (P < 0.05, 2-tailed t-test) between the V (%) value of B. mucronatus and that of B. xylophilus in this study and it is well known that these two species are morphologically too similar to distinguish each other except the shape of tail terminus (Braasch, 2008; Mamiya and Enda, 1979).
Morphometric data of male Bursaphelenchus mucronatus collected in South Korea. Mean ± SD are shown with the range of measurement in parentheses

Morphometric data of female Bursaphelenchus mucronatus collected in South Korea. Mean ± SD are shown with the range of measurement in parentheses
The morphometric characters of male B. xylophilus such as the c ratio and spicule length were affected by locational area rather than host plant, while that of female B. xylophilus such as the b ratio was affected by host plant rather than locational area. The body length of B. xylophilus was affected by locational area, host plant, or both. In this study, it was confirmed that some morphometric characters of B. xylophilus were affected by locational area, host plant, or both and those characters showed extended range of variation. When using these characters in taxonomic studies, we should be careful of the amount of variation of those characters.